ATP Depletion and Energy Metabolism in Dormant Bacteria: Mechanisms, Therapeutic Strategies, and Clinical Implications
This article comprehensively examines the critical role of ATP depletion and metabolic reprogramming in bacterial dormancy and persistence.
ATP Depletion and Energy Metabolism in Dormant Bacteria: Mechanisms, Therapeutic Strategies, and Clinical Implications
Abstract
This article comprehensively examines the critical role of ATP depletion and metabolic reprogramming in bacterial dormancy and persistence. Targeting researchers, scientists, and drug development professionals, it synthesizes foundational science on how reduced intracellular ATP triggers a dormant state, enabling pathogens like Staphylococcus aureus and Escherichia coli to tolerate antibiotics. The content explores methodological advances for studying bacterial energetics, innovative therapeutic strategies that exploit or reverse metabolic dormancy to combat persistent infections, and comparative analyses of mechanisms across species. By integrating cutting-edge research on bioenergetic stress, protein aggregation, and metabolite-driven resuscitation, this review provides a roadmap for developing novel anti-persister therapies that address the global challenge of antibiotic treatment failure.
The Energetic Basis of Bacterial Dormancy: From ATP Depletion to Persister Formation
Bacterial persisters represent a transiently antibiotic-tolerant subpopulation that significantly contributes to chronic and recurrent infections, posing a substantial challenge in clinical therapeutics. Unlike genetic resistance, persistence operates through phenotypic switching into a dormant or slow-growing state without genetic alterations. This technical review examines the defining characteristics of bacterial persisters, with particular emphasis on ATP depletion and energy metabolism as central mechanisms in dormancy establishment and maintenance. We synthesize current molecular understanding of persistence mechanisms, detail standardized experimental methodologies for persister identification and characterization, and provide visualizations of key signaling pathways. The comprehensive analysis presented herein aims to equip researchers and drug development professionals with the foundational knowledge and technical frameworks necessary to advance therapeutic strategies against persistent bacterial infections.
The concept of bacterial persistence was first formally described by Joseph Bigger in 1944, when he observed that a small subpopulation of Staphylococci survived penicillin exposure despite genetic susceptibility to the antibiotic [1] [2]. These surviving cells, which he termed "persisters," were not resistant mutants but exhibited transient tolerance that disappeared once antibiotics were removed [3]. This discovery established the fundamental distinction between genetic resistance mechanisms and the phenotypic tolerance that characterizes bacterial persistence.
In contemporary microbiology, persisters are defined as genetically susceptible bacterial cells that enter a transient, non-growing, or slow-growing state, enabling survival during exposure to lethal concentrations of antibiotics, with the ability to resume growth once the antibiotic pressure is removed [1] [4] [5]. This phenomenon is now recognized as a major contributor to treatment failure in chronic and recurrent infections, including those caused by Mycobacterium tuberculosis, Pseudomonas aeruginosa, and Staphylococcus aureus [1] [4].
The clinical significance of persisters extends beyond their role in prolonging infections. Evidence suggests that persister cells may serve as a reservoir for the evolution of genetic resistance by surviving antibiotic treatment and accumulating resistance mutations during subsequent growth phases [4] [6]. This dual threat positions persister formation as a critical focus for antimicrobial development and treatment optimization.
Defining Characteristics: Persistence vs. Resistance vs. Tolerance
Understanding bacterial persistence requires clear differentiation from related concepts of antibiotic resistance and tolerance. The table below summarizes the key distinguishing characteristics:
Table 1: Comparative Analysis of Bacterial Survival Mechanisms
| Characteristic | Susceptible Cells | Persister Cells | Tolerant Cells | Resistant Cells |
|---|---|---|---|---|
| Minimum Inhibitory Concentration (MIC) | Normal MIC | Normal MIC [6] | Normal MIC | Elevated MIC [6] |
| Genetic Basis | No resistance mutations | No resistance mutations; phenotypic variation [5] | No resistance mutations; can be phenotypic or genetic | Genetic mutations or acquired resistance genes [6] |
| Population Distribution | Majority of population | Small subpopulation (typically 0.001%-1%) [1] [7] | Can affect entire population | Selected mutants within population |
| Killing Kinetics | Rapid killing | Biphasic killing with a subpopulation dying slowly [4] [6] | Uniformly slow killing across population | Growth possible at MIC |
| Heritability | Not applicable | Non-heritable phenotype [5] [6] | Can be heritable or non-heritable | Heritable |
| Metabolic State | Active metabolism | Reduced metabolic activity, dormant, or slow-growing [1] [8] | Reduced growth rate | Typically active metabolism |
A key identifying feature of persister populations is the biphasic killing curve observed during time-kill assays, where the majority of cells die rapidly while a small subpopulation survives prolonged antibiotic exposure [4] [6]. This reflects the phenotypic heterogeneity within an isogenic population, where persisters pre-exist rather than being induced by antibiotic exposure.
It is important to distinguish between antibiotic tolerance (a property of entire populations characterized by slow killing) and persistence (a property of small subpopulations within a larger susceptible population) [6]. While all persisters are tolerant, not all tolerant cells are persisters; tolerance refers to the slow death kinetics of a population, while persistence specifically describes the biphasic killing pattern.
Molecular Mechanisms of Persister Formation
Central Role of ATP Depletion and Metabolic Dormancy
ATP depletion represents a fundamental mechanism in persister formation across bacterial species. Reduced intracellular ATP levels trigger a shift to a dormant-like state that enhances antibiotic tolerance by limiting energy-dependent processes essential for antibiotic-mediated killing [8].
Table 2: Experimental Evidence Linking ATP Depletion to Persistence
| Experimental Finding | Bacterial Species | Implication for Persistence |
|---|---|---|
| Quercetin treatment depletes ATP in dose-dependent manner (22-36% reduction) [8] | Staphylococcus aureus | Direct link between ATP depletion and persistence established |
| ATP depletion restricts energy-intensive processes (transcription, translation, cell wall synthesis) [8] | Multiple species | Fundamental mechanism for antibiotic tolerance |
| Lower ATP levels in persisters compared to vegetative cells [8] | Multiple species | Metabolic signature of persistent state |
| Quercetin pre-treatment increases persister numbers (up to 217-fold with tobramycin) [8] | Staphylococcus aureus | ATP depletion induces phenotypic switch to persistence |
The connection between ATP depletion and antibiotic efficacy is particularly pronounced for aminoglycoside antibiotics like tobramycin, which rely on proton motive force and active metabolism for intracellular uptake and efficacy [8]. The observed 217-fold increase in persister cells with tobramycin following quercetin-induced ATP depletion demonstrates the critical role of energy metabolism in antibiotic susceptibility [8].
Key Signaling Pathways and Molecular Triggers
Multiple interconnected molecular systems contribute to persister formation through their influence on bacterial metabolism and growth status:
Pathway Integration and Cross-Talk: These systems do not operate in isolation. For example, the HipA toxin of the HipAB TA system phosphorylates glutamyl-tRNA synthetase, triggering RelA-mediated (p)ppGpp synthesis and stringent response activation [5] [6]. Similarly, type I TA systems like TisB/istR and hokB/sokB can disrupt proton motive force, leading to reduced ATP levels and metabolic dormancy [6]. This interconnected network ensures robust persistence induction under diverse stress conditions.
Experimental Protocols for Persister Research
Standardized Persister Detection and Quantification
Reliable identification and quantification of persister cells requires carefully controlled methodologies. The following protocol outlines standard approaches for persister enumeration:
Critical Protocol Parameters:
- Antibiotic Concentration: Typically 5-100× MIC to ensure lethal exposure while avoiding concentration-dependent off-target effects [4] [9].
- Treatment Duration: Varies by antibiotic mechanism; 3-24 hours to capture biphasic killing [4].
- Culture Conditions: Both exponential and stationary phase cultures should be tested as persister frequencies differ significantly [1] [9].
- Confirmation of Susceptibility: Surviving colonies must be recultured and tested for unchanged MIC to confirm phenotypic rather than genetic resistance [6].
Single-Cell Analysis Using Microfluidic Devices
Advanced microfluidic approaches enable real-time observation of persister cell dynamics at single-cell resolution:
MCMA (Membrane-Covered Microchamber Array) Workflow:
- Device Fabrication: Etch 0.8-μm deep microchambers on glass coverslip [9].
- Cell Loading: Enclose bacterial cells in microchambers using cellulose semipermeable membrane [9].
- Medium Control: Manipulate environmental conditions via medium flow above membrane (complete exchange within 5 minutes) [9].
- Time-Lapse Imaging: Monitor individual cell growth, division, and antibiotic response [9].
- Phenotype Tracking: Correlate pre-exposure growth history with survival outcomes [9].
Key Insights from Single-Cell Studies:
- Persisters can originate from both growing and non-growing subpopulations depending on antibiotic class [9].
- Heterogeneous survival dynamics exist, including continuous growth with L-form-like morphologies, responsive growth arrest, or post-exposure filamentation [9].
- For β-lactams like ampicillin, most persisters in exponential phase are growing before treatment, while stationary phase yields more non-growing persisters [9].
- For fluoroquinolones like ciprofloxacin, nearly all persisters derive from growing cells regardless of culture phase [9].
Research Reagent Solutions Toolkit
Table 3: Essential Reagents and Materials for Persistence Research
| Reagent/Material | Specification/Function | Application Examples |
|---|---|---|
| Microfluidic Devices | MCMA (Membrane-Covered Microchamber Array) with 0.8-μm deep chambers [9] | Single-cell analysis of persister dynamics and lineage tracing |
| ATP Measurement Kits | Luciferase-based assays for intracellular ATP quantification [8] | Monitoring metabolic dormancy and energy depletion |
| Metabolic Stressors | Quercetin (1-10 mM in DMSO) - induces ATP depletion [8] | Experimental induction of persistence via metabolic modulation |
| Antibiotic Panels | β-lactams (ampicillin), Fluoroquinolones (ciprofloxacin), Aminoglycosides (tobramycin) [8] [9] | Persister profiling across antibiotic classes with different mechanisms |
| Bacterial Strains | Wild-type (MG1655 E. coli) and high-persistence mutants (hipA7) [1] [5] | Comparative studies of persistence mechanisms and frequencies |
| Viability Stains | Membrane integrity dyes (propidium iodide) and metabolic activity probes (CFDA) | Differentiating between dormant, dead, and active cells |
| Gene Expression Reporters | Fluorescent protein fusions (GFP, mCherry) for stress response promoters [9] | Monitoring transcriptional activation of persistence pathways |
Bacterial persistence represents a distinct phenotypic state characterized by transient antibiotic tolerance without genetic alteration, fundamentally differentiated from genetic resistance by its non-heritable nature and normal MIC. The dormancy associated with persistence is intimately connected to cellular energy metabolism, with ATP depletion serving as both a marker and mechanism of the persistent state. Understanding these distinctions is crucial for developing effective therapeutic strategies against chronic and recurrent infections.
The experimental frameworks and technical tools outlined in this review provide researchers with standardized approaches for investigating persistence mechanisms, particularly those related to energy metabolism. Future research directions should focus on elucidating the precise molecular connections between metabolic regulation, ATP homeostasis, and phenotypic switching to persistence, potentially identifying novel targets for anti-persister therapeutics. As evidence continues to establish the role of persisters in facilitating the development of genetic resistance, targeting these dormant populations becomes increasingly imperative for comprehensive antimicrobial stewardship.
Adenosine triphosphate (ATP) serves as a fundamental regulator of bacterial metabolism, with its depletion acting as a primary trigger for a state of metabolic quiescence and antibiotic tolerance. This whitepaper synthesizes current research demonstrating how bioenergetic stress—characterized by a reduced ATP/ADP ratio and adenylate energy charge—potentiates the formation of bacterial persister cells across pathogens including Staphylococcus aureus and Escherichia coli. The mechanisms linking ATP depletion to metabolic dormancy involve activation of the stringent response, reduced proton motive force, and impaired antibiotic uptake. Understanding these pathways provides critical insights for developing novel therapeutic strategies that target bacterial energetics to overcome antimicrobial tolerance.
Antimicrobial resistance represents a global health crisis, with persistent bacterial infections posing particularly challenging clinical problems. Unlike genetic resistance, antibiotic tolerance enables bacterial survival without altering minimum inhibitory concentrations through a non-heritable, phenotypic adaptation [10]. Central to this tolerance is the formation of persister cells—metabolically dormant subpopulations that withstand antibiotic exposure despite genetic susceptibility [8] [10].
The adenylate energy charge serves as a crucial indicator of cellular energy status, with ATP depletion representing a conserved stress response across bacterial species [11]. Research demonstrates that bioenergetic stress creates a state where ATP consumption exceeds production, significantly decreasing both the ATP/ADP ratio and overall adenylate energy charge [11]. This energy depletion triggers a systematic downshift to metabolic quiescence, characterized by reduced metabolic activity and biosynthetic processes, thereby enhancing bacterial survival against antibiotics that target active cellular functions [8] [10] [11].
Quantitative Evidence: Experimental Data Linking ATP Depletion to Persister Formation
Direct Measurements of ATP Depletion and Its Consequences
Table 1: Quantifiable Effects of Metabolic Stressors on ATP Levels and Bacterial Persistence
| Stress Inducer | Pathway Targeted | ATP Reduction | Persister Increase | Key Antibiotics Affected | Experimental Model |
|---|---|---|---|---|---|
| Quercetin (1 mM) | Electron transport chain, ATP synthase [8] | 22% [8] | 7-10 fold [8] | Oxacillin, Ciprofloxacin, Tobramycin [8] | Staphylococcus aureus [8] |
| Quercetin (10 mM) | Electron transport chain, ATP synthase [8] | 36% [8] | 26-217 fold [8] | Oxacillin, Ciprofloxacin, Tobramycin [8] | Staphylococcus aureus [8] |
| ATP synthase F1 complex (pF1) | Constitutive ATP hydrolysis [11] | Significant decrease in ATP/ADP & AEC [11] | Significant increase [11] | Ciprofloxacin, Gentamicin, Ampicillin [11] | Escherichia coli MG1655 [11] |
| NADH oxidase (pNOX) | Constitutive NADH oxidation [11] | Decreased ATP/ADP & AEC [11] | Significant increase [11] | Ciprofloxacin, Gentamicin [11] | Escherichia coli MG1655 [11] |
Table 2: Antibiotic Class Susceptibility to ATP Depletion-Induced Persistence
| Antibiotic Class | Example | Primary Mechanism | Fold-Change in Persisters with Quercetin [8] | Efficacy Against ATP-Depleted Cells |
|---|---|---|---|---|
| Cell Wall Synthesis Inhibitors | Oxacillin | Inhibits peptidoglycan cross-linking | 63-fold | Strongly reduced [8] |
| DNA Replication Inhibitors | Ciprofloxacin | Inhibits DNA gyrase and topoisomerase IV | 88-fold | Strongly reduced [8] [11] |
| Protein Synthesis Inhibitors (Aminoglycosides) | Tobramycin | Binds 30S ribosomal subunit | 217-fold | Most strongly reduced [8] |
| Protein Synthesis Inhibitors (Other) | Gentamicin | Binds 30S ribosomal subunit | Significant increase | Strongly reduced [11] |
Methodological Framework for Assessing ATP-Mediated Persistence
Experimental Protocol: Measuring Intracellular ATP Levels
- Culture Preparation: Grow bacterial cultures to mid-exponential phase in appropriate medium [8].
- Treatment Application: Apply metabolic stressor (e.g., quercetin) at varying concentrations for defined duration [8].
- ATP Extraction: Use dedicated ATP extraction buffers or boiling methods to liberate intracellular ATP.
- Quantification Assay: Employ luciferase-based ATP detection systems where luciferase catalyzes light emission proportional to ATP concentration [8].
- Normalization: Normalize ATP values to protein content or cell count for comparative analysis [8].
Experimental Protocol: Persister Cell Enumeration
- Antibiotic Exposure: Treat bacterial cultures with lethal concentrations of target antibiotics (typically 10-100× MIC) for defined periods [8] [11].
- Viable Counting: After antibiotic treatment, wash cells to remove antibiotics, serially dilute, and plate on antibiotic-free medium [8].
- Colony Forming Units (CFU) Enumeration: Count resulting colonies after incubation to determine surviving persister fractions [8] [11].
- Timing Considerations: Implement pre-treatment, co-treatment, and post-treatment paradigms to establish causality [8].
Molecular Mechanisms: Pathway Analysis of ATP-Mediated Metabolic Quiescence
Key Pathway Components and Regulatory Networks
The molecular pathway illustrates how ATP depletion initiates a cascade toward metabolic quiescence and antibiotic tolerance. Bioenergetic stress directly reduces the proton motive force, impairing energy-dependent antibiotic uptake mechanisms—particularly critical for aminoglycosides like tobramycin [8]. Concurrently, ATP depletion activates the stringent response via (p)ppGpp signaling, globally reprogramming cellular metabolism toward conservation [10] [11]. This metabolic rewiring further amplifies ROS production through enhanced respiration, creating oxidative stress that contributes to DNA damage and mutagenesis, potentially accelerating resistance evolution [11].
Research Reagent Solutions: Essential Tools for Investigating Bacterial Energetics
Table 3: Key Research Reagents for Studying ATP-Mediated Persistence
| Reagent/Category | Specific Examples | Research Application | Key Function |
|---|---|---|---|
| ATP Depletion Inducers | Quercetin [8] | Induce metabolic stress & dormancy | Inhibits electron transport chain & ATP synthase [8] |
| pF1 plasmid (ATP synthase F1 complex) [11] | Synthetic bioenergetic stress | Constitutive ATP hydrolysis [11] | |
| NADH Oxidation Systems | pNOX plasmid (NADH oxidase) [11] | Induce redox stress | Constitutive NADH oxidation [11] |
| ATP Quantification Kits | Luciferase-based assays [8] | Measure intracellular ATP | Bioluminescent detection of ATP concentration [8] |
| Metabolic Profiling | LC-MS/MS [11] | Comprehensive metabolomics | Quantify ATP/ADP ratios, AEC, metabolic intermediates [11] |
| Genetic Tools | cheY/flhD deletion mutants [11] | Investigate energy allocation | Reduce ATP consumption from motility [11] |
Methodological Framework: Experimental Approaches for Investigating Bacterial Energetics
Protocol for Inducing and Quantifying Bioenergetic Stress
Genetic Induction System for Bioenergetic Stress
- Plasmid Construction: Engineer low-copy plasmids containing constitutive expression cassettes for E. coli's soluble ATP synthase F1 complex (atpAGD; pF1) or Streptococcus pneumoniae NADH oxidase (nox; pNOX) with appropriate empty vector controls [11].
- Transformation & Validation: Introduce constructs into target bacterial strains and validate expression via RNA sequencing and proteomic analysis [11].
- Metabolic Confirmation: Verify bioenergetic stress through LC-MS/MS quantification of ATP/ADP ratios and adenylate energy charge [11].
Metabolomic Profiling of Bioenergetic Status
- Sample Preparation: Culture bacterial cells to exponential phase, apply treatments, and rapidly quench metabolism using cold methanol or similar methods [11].
- Metabolite Extraction: Implement dual-phase extraction to capture polar and non-polar metabolites [11].
- LC-MS/MS Analysis: Utilize reverse-phase or HILIC chromatography coupled to tandem mass spectrometry for targeted quantification of adenine nucleotides and related metabolites [11].
- Data Analysis: Calculate ATP/ADP ratios, adenylate energy charge ([ATP]+0.5[ADP])/([ATP]+[ADP]+[AMP]), and NADH/NAD+ ratios from absolute quantifications [11].
Advanced Techniques for Persister Cell Characterization
Metabolic Flux Analysis in Persister Cells
- Isolation Techniques: Implement fluorescence-activated cell sorting (FACS) based on dye retention or reporter systems to isolate persister subpopulations [10].
- Stable Isotope Tracing: Utilize 13C-labeled carbon sources (glucose, glycerol) to track metabolic activity in persister versus normal cells [10].
- Respiration Measurements: Employ extracellular flux analyzers to simultaneously measure oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) in real-time [11].
ATP serves as a master regulator connecting bioenergetic status to bacterial phenotypic states, with depletion driving a transition to metabolic quiescence and antibiotic tolerance. The experimental evidence demonstrates that diverse stressors—from flavonoid compounds like quercetin to genetic induction of ATP hydrolysis—converge on reducing ATP/ADP ratios and adenylate energy charge, thereby promoting persister formation across bacterial species and antibiotic classes [8] [11]. The mechanistic insights into this process reveal multiple intervention points for therapeutic development, particularly strategies that manipulate bacterial metabolic states to resensitize persisters to conventional antibiotics [10].
Future research directions should focus on translating these fundamental discoveries into clinical applications, including metabolite-based adjuvant therapies that reprogram persister cell metabolism, and combination treatments that prevent ATP depletion-induced tolerance. Understanding the nuanced relationship between ATP concentration, metabolic flux, and phenotypic resistance will enable more effective strategies against chronic and recurrent bacterial infections.
Bacterial dormancy represents a significant challenge in clinical medicine, contributing to chronic and relapsing infections that are difficult to eradicate with conventional antibiotics. At the heart of this phenomenon lies a coordinated physiological shutdown driven by specific molecular triggers. This technical guide examines the core pathways that induce and regulate bacterial dormancy, with particular emphasis on their convergence toward ATP depletion and energy metabolism disruption. Understanding these interconnected mechanisms—the stringent response, toxin-antitoxin (TA) modules, and related stress pathways—provides critical insights for developing novel therapeutic strategies against persistent bacterial infections.
Molecular Mechanisms of Persister Formation and Survival
Bacterial persisters are non-growing or slow-growing phenotypic variants that survive antibiotic exposure and other lethal stresses without genetic resistance. These dormant cells can resume growth after stress removal, contributing to infection relapse and treatment failure [12]. Multiple interconnected molecular systems regulate the entry into this dormant state, primarily through metabolic downregulation and energy depletion [10].
The Stringent Response and (p)ppGpp Signaling
The stringent response is a global adaptation mechanism activated by nutrient limitation and other stress conditions. This pathway is centrally mediated by the alarmones guanosine tetraphosphate and pentaphosphate (collectively ppGpp), which orchestrate a massive reprogramming of cellular physiology.
- Activation Triggers: The stringent response is primarily triggered by amino acid starvation and nutrient limitation. Specifically, uncharged tRNAs in the ribosomal A-site activate RelA, a key ppGpp synthase [13]. A second enzyme, SpoT, can both synthesize and hydrolyze ppGpp, shifting toward synthesis under various nutrient stresses including fatty acid starvation [13].
- Physiological Consequences: Elevated ppGpp levels lead to comprehensive transcriptional reprogramming, inhibiting stable RNA synthesis while promoting expression of stress resistance and survival genes. This signaling molecule directly reduces intracellular ATP levels, creating a state of energy depletion that characterizes dormant persister cells [10]. Through this metabolic rewiring, the stringent response induces a dormant state that enhances bacterial survival under adverse conditions.
Toxin-Antitoxin (TA) Modules
TA systems are genetic elements encoding a stable toxin that can inhibit cell growth and a labile antitoxin that neutralizes the toxin. These modules function as bacterial stress response systems that directly induce dormancy under unfavorable conditions.
- Induction of Dormancy: TA systems are activated by various environmental stresses, including antibiotic exposure, nutrient limitation, and oxidative stress [12]. When activated, toxins target essential cellular processes such as protein translation, DNA replication, and cell wall synthesis [10]. For example, the VapC21 toxin in Mycobacterium tuberculosis promotes persister cell formation against multiple antibiotics when overexpressed [14].
- Connection to Energy Metabolism: Toxin activity frequently leads to ATP depletion through inhibition of central metabolic pathways. This energy depletion reinforces the dormant state by limiting energy-intensive processes, creating a self-sustaining cycle of metabolic quiescence [10]. The formation of type II persisters throughout the exponential growth phase has been linked to stochastic activation of TA systems [7].
Protein Aggregation and Metabolic Sequestration
Recent research has revealed protein aggregation as a significant driver of bacterial dormancy, providing a direct link between protein homeostasis and metabolic shutdown.
- Aggregation Process: Under stress conditions, proteins progressively assemble into liquid-like condensates that subsequently undergo a liquid-to-solid phase transition [15]. This maturation process impacts the reversibility of dormancy, with solid aggregates being more difficult to dissolve.
- Metabolic Consequences: Aggregates selectively sequester proteins involved in energy production and central metabolism [15]. This sequestration directly depletes available enzymes for ATP generation, leading to progressively reduced energy levels. The timing of protein aggregation consistently precedes or coincides with the development of persistence across different genetic backgrounds, suggesting a causal relationship [15].
Table 1: Key Molecular Triggers of Bacterial Dormancy
| Molecular Trigger | Primary Activators | Core Effectors | Downstream Effects on Energy Metabolism |
|---|---|---|---|
| Stringent Response | Nutrient limitation, amino acid starvation, fatty acid starvation | (p)ppGpp, RelA, SpoT | Reduces intracellular ATP levels, inhibits energy-intensive processes |
| Toxin-Antitoxin Modules | Antibiotic exposure, oxidative stress, DNA damage | Toxins (e.g., VapC21, TisB), Antitoxins | Depletes ATP through inhibition of metabolic pathways |
| Protein Aggregation | Nutrient starvation, oxidative stress | Protein condensates, chaperones (DnaK, ClpB) | Sequesters enzymes involved in energy production |
| SOS Response | DNA damage, antibiotic stress | LexA, RecA | Induces TisB toxin expression, inhibits ATP synthesis |
Quantitative Experimental Data on ATP Depletion and Persister Formation
Recent studies have provided quantitative evidence linking molecular triggers to ATP depletion and persister formation. The following table summarizes key experimental findings from the literature.
Table 2: Quantitative Data on Molecular Triggers and Persister Formation
| Experimental System | Induction Method | ATP Reduction | Persister Increase | Key Measurements |
|---|---|---|---|---|
| S. aureus + Quercetin [8] | Quercetin treatment (dose-dependent) | 22% (1 mM), 36% (10 mM) | 63-217 fold (varies by antibiotic) | intracellular ATP levels, CFU after antibiotic exposure |
| E. coli protein aggregation [15] | Starvation-induced aggregation | Correlation with aggregate formation | Preceded by aggregation | IbpA-msfGFP foci, phase-bright aggregates, CFU |
| M. tuberculosis VapC21 [14] | Toxin overexpression | Not quantified | Increased tolerance to amikacin, streptomycin, ethambutol | CFU after antibiotic exposure |
| E. coli LplA salvage pathway [16] | Enhanced lipoylation | Not directly measured | Not applicable | PDH/OGDH activity, oxidative phosphorylation |
Experimental Protocol: Quercetin-Induced ATP Depletion and Persistence
The following detailed methodology outlines the approach used to investigate quercetin-induced metabolic stress in S. aureus [8]:
Bacterial Strains and Culture Conditions:
- Use Staphylococcus aureus reference strains (e.g., ATCC strains) and relevant clinical isolates.
- Culture bacteria in appropriate liquid media (e.g., Tryptic Soy Broth or Mueller-Hinton Broth) overnight at 37°C with shaking.
Quercetin Treatment and ATP Measurement:
- Prepare fresh quercetin solutions in suitable solvent (e.g., DMSO) with final concentrations typically ranging from 0.1 mM to 10 mM.
- Dilute overnight cultures to approximately 10^6 CFU/mL in fresh media containing varying quercetin concentrations.
- Incubate treated cultures for predetermined times (e.g., 2-6 hours) at 37°C with shaking.
- Measure intracellular ATP levels using commercially available bioluminescence assay kits based on luciferase activity.
- Normalize ATP readings to bacterial cell counts or protein content.
Persister Cell Assay:
- After quercetin pretreatment, expose cultures to lethal concentrations of antibiotics (e.g., oxacillin, ciprofloxacin, tobramycin) for 24 hours.
- Determine viable counts by serially diluting and plating on drug-free solid media.
- Calculate persister frequency as CFU/mL after antibiotic treatment divided by initial CFU/mL.
- Include controls without quercetin pretreatment and without antibiotic exposure.
Timing Experiments:
- For pre-treatment studies, expose bacteria to quercetin for 1 hour before antibiotic addition.
- For co-treatment studies, add quercetin and antibiotics simultaneously.
- For post-treatment studies, add quercetin after antibiotic exposure to assess resuscitation effects.
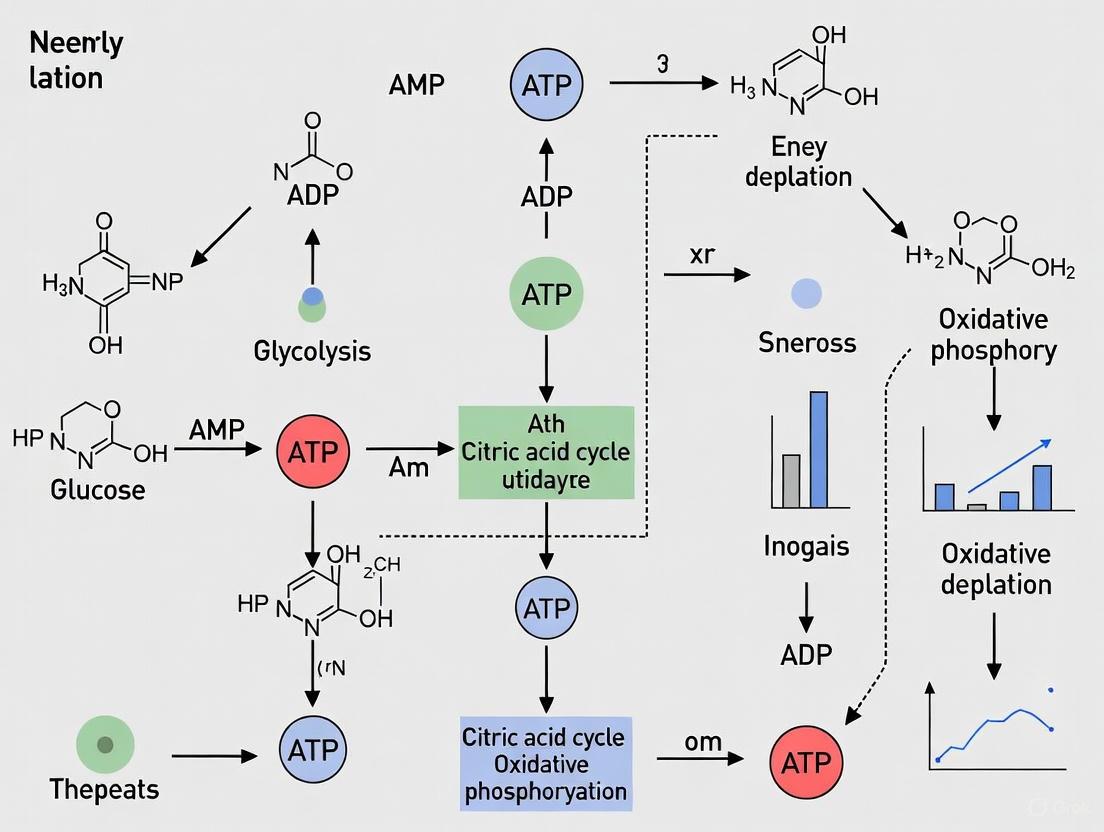
Visualization of Molecular Pathways
The diagram below illustrates the interconnected pathways through which stringent response, TA modules, and stress responses converge to induce ATP depletion and bacterial dormancy.
The Scientist's Toolkit: Research Reagents and Essential Materials
This section catalogues critical reagents and methodologies employed in studying molecular triggers of bacterial dormancy, providing researchers with practical resources for experimental design.
Table 3: Essential Research Reagents for Investigating Dormancy Triggers
| Reagent / Method | Specific Examples | Research Application | Key Functions |
|---|---|---|---|
| ATP Measurement Kits | BacTiter-Glo, ATP bioluminescence assays | Quantifying intracellular ATP levels | Measures energy depletion in persister cells |
| Fluorescent Protein Reporters | IbpA-msfGFP [15] | Tracking protein aggregation | Labels early-stage protein aggregates |
| Genetic Mutants | ΔrelA, ΔspoT, hipA, ΔrpoS [13] [15] | Dissecting pathway contributions | Tests necessity of specific genes in persistence |
| Inducible Expression Systems | Tetracycline-inducible promoters | Controlled toxin expression | Enables precise timing of dormancy induction |
| Metabolite Supplements | Lipoic acid, pyruvate, mannitol [10] [16] | Metabolic resuscitation studies | Reverses dormancy by boosting energy metabolism |
| Chemical Inducers | Quercetin [8] | Inducing metabolic stress | Triggers ATP depletion and persistence |
| Antibiotic Selection | Oxacillin, ciprofloxacin, tobramycin [8] | Persister isolation and quantification | Kills normal cells while sparing persisters |
Therapeutic Implications and Future Perspectives
The molecular understanding of bacterial persistence mechanisms opens promising avenues for therapeutic development. Current research focuses on metabolite-driven resuscitation and anti-dormancy compounds that target the specific mechanisms maintaining bacterial dormancy.
Metabolite-Based Adjuvants: Exogenous metabolites such as pyruvate, mannitol, and lipoic acid can restore metabolic activity in dormant cells [10]. This "wake and kill" approach resensitizes persisters to conventional antibiotics by reactivating central energy metabolism and restoring proton motive force [10]. For instance, Allison et al. demonstrated that metabolites enhance aminoglycoside uptake and bactericidal activity by restoring proton motive force in vitro and in animal models [10].
Protein Aggregation Interventions: Targeting the liquid-to-solid transition of protein aggregates may prevent the development of deeply dormant VBNC cells [15]. Chaperones like DnaK and ClpB that facilitate aggregate dissolution represent potential therapeutic targets to promote resuscitation of dormant cells [15].
Combination Therapies: The future of combating persistent infections likely lies in antibiotic-adjuvant combinations that simultaneously target active populations and dormant persisters. As quercetin studies demonstrate, careful timing is essential, as some compounds can both inhibit growth and promote persistence depending on administration protocol [8].
The ongoing challenge involves translating these mechanistic insights into effective clinical treatments that can overcome the antibiotic tolerance conferred by bacterial dormancy.
Metabolic reprogramming, a established hallmark of cancer, is increasingly recognized as a critical adaptive mechanism in bacterial pathogenesis, particularly in the context of dormancy and persistence [17] [18]. This rewiring of core metabolic pathways enables bacterial cells to survive antibiotic treatment and other environmental stresses by drastically reducing their metabolic activity and entering a dormant state [8]. At the heart of this phenomenon lies the reprogramming of central carbon metabolism and respiration, leading to significant ATP depletion—a key characteristic of bacterial persister cells [8]. This whitepaper provides an in-depth technical analysis of the principal metabolic hallmarks involved in this reprogramming, with a specific focus on its implications for ATP biosynthesis and energy metabolism. It is intended to equip researchers and drug development professionals with a detailed framework and methodologies for investigating these processes, ultimately informing the development of novel therapeutic strategies against persistent bacterial infections.
Core Hallmarks of Metabolic Reprogramming
Metabolic reprogramming in dormant bacteria encompasses several interconnected alterations to core biochemical pathways. These hallmarks are orchestrated to reduce energy expenditure, maintain redox balance, and support survival under duress, with ATP depletion being a consistent outcome [8] [18].
Table 1: Core Hallmarks of Metabolic Reprogramming in Bacterial Dormancy and Persistence
| Hallmark | Key Alterations | Functional Consequences | Link to ATP Depletion & Dormancy |
|---|---|---|---|
| Increased Glycolysis & Lactic Acid Production | Upregulation of glucose transporters and glycolytic enzymes; diversion of pyruvate to lactate via LDH [18]. | Rapid but inefficient ATP yield (2 ATP/glucose); generation of biosynthetic precursors [18]. | Low ATP yield per glucose molecule contributes to overall energy deficit; lactate accumulation can inhibit growth [8] [18]. |
| Glutaminolysis | Increased uptake of glutamine via transporters (e.g., ASCT2); conversion to glutamate and TCA cycle intermediates via GLS1 [17] [18]. | Replenishes TCA cycle (anaplerosis); provides nitrogen for biosynthesis [17] [18]. | Can support limited ATP production via TCA cycle/OXPHOS, but is often downregulated in dormancy [8]. |
| Mitochondrial & Respiratory Changes | Shift from oxidative phosphorylation (OXPHOS) to aerobic glycolysis; downregulation of electron transport chain activity [18]. | Reduced mitochondrial ATP generation; altered reactive oxygen species (ROS) signaling [8] [18]. | Direct cause of severe ATP depletion; primary mechanism for reducing energy metabolism in dormant cells [8]. |
| Pentose Phosphate Pathway (PPP) Activation | Upregulation of G6PD; generation of ribose-5-phosphate and NADPH [18]. | Supports nucleotide synthesis; maintains redox balance via glutathione system [18]. | Not a direct ATP source, but crucial for survival in a non-proliferative state by mitigating oxidative stress [8]. |
The interplay of these pathways results in a profound reprogramming of central carbon metabolism, where carbon flux is redirected away from high-yield energy production in the mitochondria towards pathways that support stress tolerance and the maintenance of basic cellular functions despite a low-energy state [17] [8] [18].
Quantitative Profiling of Metabolic Changes
Advanced metabolomics technologies are essential for quantifying the spatial and temporal dynamics of these metabolic hallmarks. A recent, robust methodology employs Mass Spectrometry Imaging (MSI) with uniform 13C-labelled yeast extracts as internal standards for pixel-wise normalization, enabling highly accurate quantification of over 200 metabolic features in tissue samples [19].
Table 2: Key Metabolite Changes in Metabolic Reprogramming Linked to Stress/Dormancy
| Metabolite / Pathway | Measurable Change | Quantitative Method | Biological Interpretation |
|---|---|---|---|
| Intracellular ATP | Significant decrease (e.g., 22-36% reduction with quercetin stress) [8]. | Luciferase-based ATP assay [8]. | Marker of metabolic dormancy and antibiotic tolerance [8]. |
| Glutamine/Glutamate | Depletion in infected host cells (e.g., by S. aureus, Legionella) [17]. | LC-MS/MS with IS normalization [19]. | Indicates bacterial consumption for energy/nitrogen; host immune modulation [17]. |
| Lactate | Increase (Warburg effect) [18]. | MALDI-MSI with IS [19]. | Sign of glycolytic flux increase and shift away from OXPHOS [18]. |
| GSH/GSSG Ratio | Decrease (e.g., in infarct core) [19]. | Quantitative MSI [19]. | Indicator of oxidative stress levels, common in metabolic stress responses [19]. |
| UDP-GlcNAc / Linoleate | Decreased precursor pools [19]. | Quantitative MSI [19]. | Suggests sustained vulnerability and altered biosynthesis long-term [19]. |
Detailed Experimental Protocol: Spatial Quantitative Metabolomics
Method: Matrix-Assisted Laser Desorption/Ionization (MALDI) Mass Spectrometry Imaging with 13C-Labelled Internal Standards [19].
Workflow Summary:
Key Steps:
- Tissue Preparation: Flash-freeze tissue samples (e.g., infected host tissue) in liquid N₂. Cryo-section into thin slices (e.g., 10-12 µm) and thaw-mount onto glass slides [19].
- Internal Standard Application: Homogeneously spray a solution of uniformly 13C-labelled yeast extract across the entire tissue section using a robotic sprayer. This provides a known quantity of a vast array of metabolite standards for every pixel [19].
- Matrix Application: Apply the matrix, N-(1-naphthyl) ethylenediamine dihydrochloride (NEDC), via spraying to facilitate desorption and ionization [19].
- MALDI-MSI Acquisition: Analyze the slides using a MALDI mass spectrometer (e.g., TimsTOF flex) in negative or positive ion mode. The instrument acquires a full mass spectrum at every predefined (x,y) coordinate, creating a spatial map of metabolite abundances [19].
- Data Processing:
- Generate a list of both endogenous metabolites and their corresponding 13C-labelled IS peaks.
- For each metabolite, normalize the intensity of the endogenous peak to the intensity of its 13C-IS peak in every pixel. This corrects for spatial variations in ion suppression (matrix effect) [19].
- Compare normalized abundances across regions of interest (e.g., infected vs. non-infected areas, core vs. periphery of a bacterial microcolony).
This method is superior to traditional normalization (e.g., Total Ion Count) as it accounts for the localized matrix effect, which can vary dramatically across different tissue structures [19].
Experimental Models for Studying ATP Depletion and Dormancy
Research into metabolic reprogramming and bacterial persistence relies on well-established in vitro and in vivo models.
In Vitro Model: Quercetin-Induced ATP Depletion inStaphylococcus aureus
Objective: To investigate how metabolic stressor-induced ATP depletion promotes bacterial dormancy and antibiotic tolerance [8].
Protocol Summary:
Detailed Methodology [8]:
- Bacterial Culture and Treatment: Grow S. aureus to mid-log phase in a suitable broth. Treat cultures with varying concentrations of quercetin (e.g., 1 mM and 10 mM) for a defined period.
- ATP Measurement: Lysate bacterial cells and measure intracellular ATP levels using a luciferase-based assay. Luciferase emits light proportional to ATP concentration, allowing for quantification. Expect a dose-dependent decrease (e.g., 22-36% reduction) [8].
- Persister Cell Assay:
- Pre-treatment: Expose bacteria to quercetin for 1 hour before adding antibiotics.
- Co-treatment: Add quercetin and antibiotics simultaneously.
- Use antibiotics targeting different processes: oxacillin (cell wall), ciprofloxacin (DNA), tobramycin (protein synthesis) [8].
- Viability Assessment: After 24 hours of antibiotic exposure, serially dilute the cultures and plate them on solid agar to determine the number of colony-forming units (CFUs). Persister cells are enumerated as the surviving CFUs that can resuscitate after antibiotic removal [8].
- Key Outcome: Quercetin pre-treatment, which induces ATP depletion, leads to a significant increase in persister cell counts (e.g., 32-fold with 10 mM quercetin and oxacillin), demonstrating a direct link between metabolic stress, ATP depletion, and antibiotic tolerance [8].
Research Reagent Solutions
The following reagents are essential for studying metabolic reprogramming and bacterial dormancy.
Table 3: Essential Research Reagents and Materials
| Reagent/Material | Specific Example / Catalog Number | Function in Experimental Protocol |
|---|---|---|
| U-13C Labelled Yeast Extract | Custom-prepared or commercial isotopically labelled yeast extract [19]. | Serves as a comprehensive source of internal standards for quantitative MSI, enabling pixel-wise normalization [19]. |
| Quercetin | Sigma-Aldrich Q4951 or equivalent [8]. | A metabolic stressor used to induce ATP depletion and model the formation of dormant bacterial persister cells [8]. |
| BacTiter-Glo Assay | Promega (G8231) or similar [8]. | Luciferase-based kit for sensitive quantification of intracellular ATP levels in bacterial cultures [8]. |
| NEDC Matrix | N-(1-Naphthyl)ethylenediamine dihydrochloride [19]. | Matrix for MALDI-MSI analysis, enabling desorption/ionization of metabolites from tissue surfaces [19]. |
| Specific Antibiotics | Oxacillin, Ciprofloxacin, Tobramycin [8]. | Used in persister cell assays to apply selective pressure and quantify tolerant bacterial subpopulations [8]. |
| MALDI Mass Spectrometer | TimsTOF flex (Bruker) or equivalent [19]. | High-resolution mass spectrometer for spatial metabolomics imaging, capable of mapping metabolite distributions in tissue samples [19]. |
Integrated Pathway Diagram of Metabolic Hallmarks
The diagram below synthesizes the core metabolic hallmarks and their interconnection, leading to ATP depletion and bacterial dormancy.
Bacterial phenotypic heterogeneity describes the phenomenon where genetically identical cells within a clonal population exhibit diverse physiological states, leading to differential survival outcomes under antibiotic exposure. This heterogeneity gives rise to a spectrum of bacterial persister cells—non-growing or slow-growing variants that survive antibiotic treatment despite genetic susceptibility to the drugs. These persisters are now recognized as a primary cause of chronic and relapsing infections and represent a significant challenge in clinical management of bacterial diseases [12]. The spectrum of persistence states ranges from "shallow" persisters, which exhibit modest tolerance and can resume growth relatively quickly after antibiotic removal, to "deep" persisters, which exist in a state of profound metabolic dormancy and require extended recovery periods [12].
Critically, this phenotypic heterogeneity operates as an evolutionary bet-hedging strategy, allowing bacterial populations to survive unpredictable environmental stresses, including antibiotic exposure [20]. The formation and survival of persister cells are intimately connected to energy metabolism and ATP depletion, with recent research revealing that deep persistence states are characterized by dramatic reductions in metabolic activity and energy production. This metabolic dormancy presents a formidable barrier to conventional antibiotic treatments, as most clinically used antibiotics target active cellular processes [10] [12]. Understanding the molecular mechanisms governing this continuum from shallow to deep persistence is therefore essential for developing novel therapeutic approaches to eradicate persistent infections.
Molecular Mechanisms Governing Persistence Heterogeneity
Metabolic Regulation and Energy Depletion
The transition from shallow to deep persistence states follows a continuum of progressively reduced metabolic activity and energy depletion. In shallow persisters, metabolic activity is diminished but not completely halted, whereas deep persisters experience severe ATP depletion and enter a state of metabolic quiescence that renders them highly tolerant to antibiotics [12]. This relationship between metabolic state and persistence level is not merely correlative but causal; experimental evidence demonstrates that artificially modulating metabolic activity can directly influence a cell's position along the persistence spectrum.
Central to this metabolic regulation is the stringent response mediated by the signaling molecule (p)ppGpp, which orchestrates a comprehensive reprogramming of cellular physiology in response to nutrient limitation and other stresses. This response redirects cellular resources away from growth and division toward maintenance and survival pathways, effectively tuning the cell's metabolic activity to match environmental conditions [10]. The implementation of this response varies significantly between individual cells, creating a population with heterogeneous metabolic states and consequently heterogeneous levels of antibiotic tolerance.
Table 1: Characteristics of Shallow vs. Deep Persister Cells
| Characteristic | Shallow Persisters | Deep Persisters |
|---|---|---|
| Metabolic Activity | Moderately reduced | Severely reduced or undetectable |
| ATP Levels | Partially depleted | Severely depleted |
| Recovery Time | Short (hours) | Extended (days to weeks) |
| Protein Synthesis | Low but detectable | Nearly absent |
| Antibiotic Tolerance Level | Moderate | High |
| Primary Formation Mechanism | Type I (stress-induced) | Type II (stochastic) |
Protein Aggregation in Metabolic Shutdown
A groundbreaking mechanism linking energy depletion to persistence states was recently revealed in 2025, demonstrating that protein aggregation plays a central role in bacterial dormancy development. Research on Escherichia coli showed that under stress conditions, proteins involved in energy metabolism undergo liquid-to-solid phase transitions, forming gel-like droplets that progressively solidify [21]. This aggregation process effectively shuts down bacterial energy production by sequestering metabolic enzymes into inactive complexes, creating a self-reinforcing cycle of metabolic decline.
The timing of protein condensation correlates precisely with entry into dormancy across different E. coli strains, suggesting a conserved mechanism for persistence establishment. The maturation of protein aggregates from liquid condensates to solid structures corresponds to the transition from shallow to deep persistence states, with solid aggregates characterizing the deeply persistent VBNC (viable but non-culturable) state [21]. This discovery is particularly significant as it reveals that protein aggregation—previously associated primarily with neurodegenerative diseases—can serve a protective function in bacterial cells by facilitating metabolic shutdown under stress.
Toxin-Antitoxin Systems and Persistence Regulation
Toxin-antitoxin (TA) modules represent another key mechanism generating persistence heterogeneity through targeted inactivation of essential cellular processes. These systems consist of a stable toxin and its cognate labile antitoxin; under stress conditions, protease-mediated degradation of the antitoxin releases the toxin to act on its cellular targets [10]. Different TA systems target distinct metabolic processes: HipA inhibits translation by phosphorylating glutamyl-tRNA synthetase, while TxpA inhibits translation and induces persistence by cleaving mRNA [10].
The stochastic activation of various TA systems within individual cells creates a population with heterogeneous physiological states, effectively distributing the risk of antibiotic exposure across multiple phenotypic variants. This bet-hedging strategy ensures that at least some subpopulations will survive unpredictable environmental challenges [20]. The combined action of multiple TA systems, each responding to different stress signals and targeting different cellular processes, enables the generation of a continuum of persistence states rather than a simple binary switch between growing and non-growing states.
Single-Cell Analysis of Persistence Heterogeneity
Advanced Methodologies for Resolving Persistence States
Traditional bulk measurement techniques mask the heterogeneity within bacterial populations, necessitating single-cell approaches to resolve the spectrum of persistence states. Several advanced methodologies have been developed to quantify and characterize phenotypic heterogeneity at the single-cell level:
Microfluidic single-cell imaging: The "mother machine" microfluidic device enables continuous observation of individual cells under constant growth conditions, allowing researchers to track metabolic activity and gene expression in real-time [22]. This approach revealed that apparently stochastic heterogeneity in oxidative stress responses actually arises from deterministic cell-cell interactions and local environmental gradients [22].
Nanoscale secondary ion mass spectrometry (NanoSIMS): When coupled with stable isotope probing (SIP), this technology enables precise quantification of isotope incorporation in individual microbial cells, providing direct measurement of metabolic activity at the single-cell level [23] [24]. This approach has revealed extensive heterogeneity in substrate assimilation among genetically identical cells under identical environmental conditions.
Heterogeneity quantitation indices: To standardize measurements across studies, researchers have developed two specialized indices: the Heterogeneity Coefficient (HC) for unimodal activity distributions and the Differentiation Tendency Index (DTI) for multimodal distributions indicating distinct subpopulations [23]. These quantitative tools facilitate direct comparison of heterogeneity across different experimental conditions and bacterial species.
Table 2: Quantitative Methods for Analyzing Phenotypic Heterogeneity
| Method | Resolution | Key Measurable Parameters | Throughput | Applications |
|---|---|---|---|---|
| SIP-NanoSIMS | ~50 nm | Isotopic enrichment, elemental composition | Low (~10² cells) | Metabolic activity, substrate assimilation |
| Flow Cytometry | Single cell | Fluorescence, cell size, complexity | High (~10⁵ cells) | Gene expression, membrane potential |
| Microfluidic Microscopy | Single cell | Growth rate, division time, morphology | Medium (~10³ cells) | Temporal dynamics, cell lineages |
| Single-Cell RNA Sequencing | Single cell | Transcriptome profiles | Medium (~10⁴ cells) | Gene expression heterogeneity |
Experimental Protocol: Single-Cell Analysis of Persister Metabolism
The following detailed protocol enables investigation of metabolic heterogeneity in bacterial persisters using advanced single-cell techniques:
Procedure:
- Culture Preparation: Grow bacterial cultures to mid-exponential phase in appropriate medium. For E. coli, use LB medium at 37°C with shaking at 200 rpm until OD600 reaches 0.4-0.6.
- Antibiotic Treatment: Expose cultures to bactericidal antibiotic (e.g., 5-10× MIC of ampicillin or ciprofloxacin) for 2-4 hours to eliminate non-persister cells.
- Cell Sorting: Wash antibiotic-treated cells twice with fresh medium and sort persister cells using fluorescence-activated cell sorting (FACS). For metabolic activity-based sorting, use dyes such as 5-cyano-2,3-ditolyl tetrazolium chloride (CTC) for respiration or propidium iodide for membrane integrity.
- Stable Isotope Labeling: Resuspend sorted persisters in medium containing 13C-labeled substrates (e.g., 13C-glucose or 15N-ammonium sulfate) and incubate for 2-4 hours under appropriate growth conditions.
- NanoSIMS Sample Preparation: Fix cells with 2.5% glutaraldehyde for 30 minutes, wash with PBS, and deposit onto silicon wafers. Dehydrate through ethanol series and critical point dry.
- NanoSIMS Analysis: Acquire secondary ion images for 12C-, 13C-, 12C14N-, and 12C15N- using Cs+ primary ion beam at 1-2 pA current. Collect 10-20 frames of 256×256 pixels with 1-10 ms/pixel dwell time.
- Data Processing: Calculate 13C/12C and 15N/14N ratios for individual cells using image analysis software (e.g., OpenMIMS). Normalize ratios to standard reference materials.
- Heterogeneity Quantification: Compute Heterogeneity Coefficient (HC) for unimodal distributions: HC = (90th percentile - 10th percentile) / median. For multimodal distributions, calculate Differentiation Tendency Index (DTI) using cluster analysis.
Therapeutic Targeting of Persistence States
Metabolic Reprogramming Strategies
The intimate connection between metabolic state and antibiotic tolerance suggests promising therapeutic approaches focused on metabolic reactivation of persistent cells. The "wake and kill" strategy involves administering metabolites or other compounds that reactivate bacterial metabolism, thereby resensitizing persisters to conventional antibiotics [10]. This approach takes advantage of the fundamental principle that bactericidal antibiotic efficacy correlates strongly with cellular metabolic activity [10].
Specific metabolic interventions include:
- Mannitol addition, which enhances proton motive force and promotes aminoglycoside uptake in Pseudomonas aeruginosa biofilms [10].
- Exogenous pyruvate, which has been shown to promote gentamicin uptake and killing of antibiotic-resistant Vibrio alginolyticus [10].
- L-valine supplementation, which stimulates phagocytosis and enhances clearance of multidrug-resistant bacterial pathogens [10].
- Adenosine and guanosine, which have demonstrated ability to enhance tetracycline sensitivity of persister cells [10].
These metabolite-based approaches work by reversing the metabolic dormancy that characterizes persister cells, effectively shifting cells from deeper to shallower persistence states where they become vulnerable to antibiotic killing.
Protein Aggregation Disruption
The recent discovery of protein aggregation as a key mechanism in persistence formation suggests novel therapeutic targets for combating persistent infections. Research has identified that the chaperone protein DnaK can actively dissolve protein aggregates and reactivate bacterial metabolism, facilitating bacterial resuscitation from dormant states [21]. This finding opens possibilities for developing compounds that either enhance DnaK-mediated disaggregation or prevent the initial formation of metabolic protein aggregates.
Experimental evidence indicates that disrupting the aggregation process can prevent entry into deep persistence states, potentially keeping cells in more antibiotic-sensitive shallow persistence states. This approach is particularly promising when combined with conventional antibiotics, as it may prevent the establishment of the deeply dormant cells that are responsible for relapse infections following antibiotic treatment [21].
Research Tools and Experimental Reagents
Table 3: Essential Research Reagents for Persistence Studies
| Reagent/Cell Line | Application | Key Features/Experimental Function |
|---|---|---|
| E. coli HipA7 mutant | High-persistence model | Contains hipA7 allele resulting in 100-1000× increased persistence frequency [20] |
| Microfluidic devices (mother machine) | Single-cell dynamics | Enables long-term imaging of individual cells under controlled conditions [22] |
| 13C/15N-labeled substrates | Metabolic tracking | Allows quantification of metabolic activity via SIP-NanoSIMS [23] |
| OxyR reporter strain (PgrxA-CFP) | Oxidative stress monitoring | Reports activation of OxyR regulon in response to H2O2 stress [22] |
| DnaK expression plasmids | Protein aggregation studies | Enables manipulation of chaperone activity to test resuscitation mechanisms [21] |
| pRPL28-sfGFP reporter | Growth status monitoring | Fluorescent reporter of ribosomal protein expression indicating metabolic activity [25] |
Visualizing Key Mechanisms and Workflows
Metabolic Regulation of Persistence Formation
Single-Cell Analysis Workflow
The spectrum from shallow to deep persistence states represents a sophisticated bacterial survival strategy rooted in metabolic heterogeneity and energy depletion. The continuum of persistence states, governed by mechanisms including stringent response activation, toxin-antitoxin system operation, and protein aggregation, enables bacterial populations to withstand antibiotic therapy and other environmental stresses. Advanced single-cell technologies have been instrumental in resolving this heterogeneity, revealing that previously assumed stochastic variation often arises from deterministic processes and cell-cell interactions.
Therapeutic approaches that target the metabolic basis of persistence—particularly metabolite-driven resuscitation strategies and protein aggregation disruption—hold significant promise for addressing the clinical challenge of persistent infections. By manipulating bacterial metabolic states to shift cells from deeper to shallower persistence states, these approaches potentially resensitize persisters to conventional antibiotics. Future research focusing on the precise molecular mechanisms governing transitions along the persistence spectrum, particularly the role of energy metabolism and protein homeostasis, will be essential for developing effective therapies against chronic and relapsing bacterial infections.
Investigating Bacterial Energetics: Techniques for Quantifying and Targeting Metabolic Dormancy
The measurement of intracellular adenosine triphosphate (ATP) is a cornerstone of understanding cellular energy metabolism, a field with profound implications for microbial ecology, antibiotic development, and bioproduction. Within the context of dormant bacteria research, ATP dynamics take on a critical significance. Dormant spores, once considered physiologically inert, are now known to perform sophisticated computations about their environment while in a metabolically inactive state. Groundbreaking research has revealed that dormant spores of Bacillus subtilis can process information by using stored electrochemical energy, in the form of potassium ions, to monitor their surroundings without metabolic activity [26] [27]. This "thinking undead" state allows them to integrate fleeting environmental signals and determine the optimal time to return to life, a process analogous to neuronal integrate-and-fire mechanisms [26]. Conversely, studies on the phototrophic bacterium Rhodopseudomonas palustris demonstrate a direct link between intracellular ATP depletion and the loss of viability in growth-arrested cells under energy-depleted (dark) conditions [28]. These findings frame a central thesis: ATP depletion is a fundamental driver of the dormant state, and the maintenance of energy charge is essential for long-term survival and the capacity to resume metabolic activity. Accurate measurement of intracellular ATP is therefore not merely a technical exercise but a prerequisite for unraveling the energetic principles governing bacterial persistence.
Core Analytical Techniques for ATP Profiling
The accurate quantification of ATP, along with its related metabolites ADP and AMP, allows for the calculation of the Energy Charge (EC), a critical index of cellular viability and metabolic fitness. The Energy Charge is calculated as: EC = ( [ATP] + 0.5[ADP] ) / ( [ATP] + [ADP] + [AMP] ) [29]. This section details the primary analytical strategies employed for these measurements.
HPLC-MS/MS for Direct Quantification
Liquid Chromatography coupled with tandem Mass Spectrometry (HPLC-MS/MS) represents the gold standard for sensitive and specific quantification of intracellular nucleotides.
- Principle: This technique separates ATP, ADP, and AMP based on their retention time on a chromatographic column, followed by selective detection and quantification via mass spectrometry. This avoids the need for derivatization or enzymatic conversion, allowing for the direct and simultaneous measurement of all three adenylates [30] [29].
- Workflow: The general workflow involves quenching metabolism rapidly, extracting metabolites (often with organic solvents like methanol/acetonitrile), separating the extract via HPLC, and detecting analytes using MS/MS. A recent method developed for a nucleotide analogue demonstrates the feasibility of direct triphosphate quantification, using sophisticated chromatography to separate the target from complex cellular matrices [30].
- Advantages: High specificity, ability to profile multiple metabolites simultaneously, and excellent sensitivity suitable for low biomass samples like dormant bacteria.
Genetically Encoded Biosensors for Real-Time Dynamics
For monitoring real-time ATP dynamics in living cells, genetically encoded biosensors offer an unparalleled approach.
- Principle: These are engineered fluorescent proteins that change their fluorescence properties upon binding ATP. A prominent example is the iATPsnFR1.1 sensor, which contains a circularly permuted super-folder green fluorescent protein (cp-sfGFP) integrated into the ATP-binding epsilon subunit of the F0-F1 ATP synthase. ATP binding induces a conformational change that enhances green fluorescence [31].
- Implementation: To control for variations in sensor expression, the sensor is often fused to a reference fluorescent protein like mCherry. The ratio of GFP to mCherry fluorescence provides a ratiometric measure of intracellular ATP concentration, which can be tracked in real-time across different growth phases [31].
- Application: This method was instrumental in discovering transient ATP accumulation during the transition from exponential to stationary phase in E. coli and Pseudomonas putida, a phenomenon linked to an imbalance between ATP production and consumption during growth slowdown [31].
Luciferase-Based Assays
The luciferase assay is a classical and widely used method for ATP quantification.
- Principle: It relies on the firefly luciferase enzyme, which catalyzes a light-emitting reaction in the presence of ATP, luciferin, and oxygen. The intensity of the emitted light is directly proportional to the ATP concentration in the sample.
- Role in Validation: While excellent for bulk measurements, it is often used to validate readings from other methods, such as ATP biosensors, confirming the accuracy of the dynamic profiles observed [31].
Table 1: Comparison of Primary ATP Quantification Methods
| Method | Principle | Key Advantage | Key Limitation | Suitability for Dormancy Research |
|---|---|---|---|---|
| HPLC-MS/MS | Chromatographic separation & mass spectrometric detection | High specificity & multi-analyte profiling | Requires cell lysis; complex setup | Excellent for absolute quantification & energy charge calculation |
| Genetically Encoded Biosensors | Fluorescence change upon ATP binding | Real-time, non-destructive monitoring in live cells | Requires genetic manipulation | Ideal for kinetic studies of ATP fluctuations |
| Luciferase Assay | Light emission from luciferase-ATP reaction | Highly sensitive & commercially accessible | Requires cell lysis; single time-point measurement | Good for validating viability & bulk ATP levels |
Experimental Protocol: Intracellular ATP Quantification via HPLC-MS/MS
The following protocol, adapted from modern analytical approaches, details the steps for direct quantification of adenylates from bacterial cell pellets, such as those from dormant cultures [30] [29].
Reagents and Equipment
- Chemicals: ATP, ADP, AMP analytical standards (purity >95%), HPLC-grade acetonitrile (ACN) and methanol (MeOH), acetic acid, ammonium acetate.
- Internal Standard: A stable isotope-labeled analogue, such as 2H6-tenofovir diphosphate, is recommended [30].
- Equipment: HPLC system coupled to a triple-quadrupole mass spectrometer (MS/MS), analytical column (e.g., HILIC or reversed-phase), refrigerated centrifuge, sonicator, vacuum concentrator.
Sample Preparation Workflow
Diagram 1: ATP Analysis Sample Prep
- Culture Harvesting and Quenching: Rapidly separate bacterial cells from their medium via filtration or fast centrifugation. Immediately quench metabolism by submerging the cell pellet in liquid nitrogen or cold quenching solutions (-40°C methanol) to freeze the metabolic state instantaneously.
- Intracellular Metabolite Extraction:
- Resuspend the cell pellet in a pre-chilled extraction solvent, typically a 70:30 (v/v) mixture of methanol and water [30].
- Perform repeated freeze-thaw cycles or use sonication on ice to ensure complete cell lysis and metabolite release.
- Centrifuge at high speed (e.g., 14,000 x g, 10 min, 4°C) to pellet cell debris.
- Collect the supernatant containing the extracted metabolites.
- Sample Clean-up and Fortification:
- Add a known amount of the internal standard (IS) to the supernatant. The IS corrects for variability during sample processing and instrument analysis [30].
- The extract may be concentrated under a vacuum and reconstituted in a solvent compatible with the HPLC-MS/MS mobile phase.
HPLC-MS/MS Analysis Conditions
- Chromatography:
- Column: HILIC or a reversed-phase column suitable for polar compounds.
- Mobile Phase: Typically a gradient between an aqueous buffer (e.g., ammonium acetate) and an organic solvent (ACN or MeOH).
- Run Time: Approximately 15 minutes per sample is a typical benchmark for good separation [30].
- Mass Spectrometry:
- Ionization: Electrospray Ionization (ESI) in negative mode.
- Detection: Multiple Reaction Monitoring (MRM). Specific precursor-to-product ion transitions are defined for each analyte and the IS for highly selective quantification.
- Example MRM Transitions: These must be optimized for the specific instrument but would target the molecular ions of ATP, ADP, and AMP.
Data Analysis and Energy Charge Calculation
- Quantification: Generate a calibration curve using the analyte standards. The ratio of the analyte's peak area to the IS peak area is plotted against concentration. This curve quantifies ATP, ADP, and AMP in the unknown samples.
- Energy Charge Calculation: Use the concentrations obtained to calculate the Energy Charge:
EC = ( [ATP] + 0.5[ADP] ) / ( [ATP] + [ADP] + [AMP] )[29].
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents for Intracellular ATP Analysis
| Reagent / Material | Function / Application | Technical Notes |
|---|---|---|
| ATP, ADP, AMP Standards | Calibration and method development | Use high-purity (>95%) standards to ensure accurate quantification [30]. |
| Stable Isotope-Labeled Internal Standard (e.g., 2H6-TDF-DP) | Normalizes analytical variability | Corrects for losses during sample prep and matrix effects in MS [30]. |
| Methanol (HPLC-MS Grade) | Metabolite extraction solvent | High-purity solvent is critical to minimize background noise in MS detection [30]. |
| Genetically Encoded ATP Biosensor (e.g., iATPsnFR1.1) | Real-time monitoring of ATP dynamics in live cells | A ratiometric sensor (fused to mCherry) is recommended for reliable results [31]. |
| Firefly Luciferase Assay Kit | Validation and bulk ATP measurement | A standard method for validating biosensor data or quickly assessing viability [31]. |
Signaling and Metabolic Pathways in Bacterial Energetics
Understanding the pathways that generate and consume ATP is essential for interpreting quantification data, especially in dormant systems.
Diagram 2: Bacterial Energy Pathways
The diagram above illustrates key energetic strategies. In dormant spores, a unique integrate-and-fire mechanism uses stored potassium ions, not metabolic ATP, to monitor the environment [26] [27]. In contrast, growth-arrested but metabolically potential cells like R. palustris rely on light to generate ATP via photophosphorylation; without this energy source, ATP depletion leads to translational shutdown and loss of viability [28]. Furthermore, recent research on Mycobacterium smegmatis has uncovered a novel pathway for energy generation from atmospheric hydrogen via the Huc hydrogenase, which directly reduces menaquinone in the respiratory chain, providing a minimal but sufficient energy source for survival in extreme, nutrient-poor environments [32]. This pathway is particularly relevant for understanding how dormant bacteria might persist for extended periods.
Data Presentation and Analysis
Effective data presentation is key to communicating findings on ATP dynamics. The table below summarizes hypothetical but representative quantitative data from different bacterial states, illustrating the power of adenylate profiling.
Table 3: Hypothetical Adenylate Profile and Energy Charge in Bacterial States
| Bacterial State / Condition | [ATP] (nmol/mg protein) | [ADP] (nmol/mg protein) | [AMP] (nmol/mg protein) | Energy Charge (EC) | Interpretation |
|---|---|---|---|---|---|
| Exponential Growth (in Glucose) | 15.0 | 5.0 | 1.0 | 0.88 | High metabolic activity & growth |
| Transition to Stationary (in Acetate) [31] | 22.5 (Peak) | 6.5 | 1.5 | 0.91 | Transient ATP surplus from growth slowdown |
| Late Stationary (in Glucose) | 7.0 | 4.0 | 2.5 | 0.67 | Reduced metabolic activity & energy charge |
| Dormant Spore (Hydrated) | < 0.1 | < 0.1 | < 0.1 | N/A | Metabolically inactive; uses ion potential [26] |
| Growth-Arrested R. palustris (Light) [28] | ~7.0 | Data not in source | Data not in source | Data not in source | Maintained viability via photophosphorylation |
| Growth-Arrested R. palustris (Dark) [28] | < 0.1 (after 25d) | Data not in source | Data not in source | Data not in source | ATP depletion correlated with 3-log loss of viability |
Bioenergetic stress represents a physiological state in bacterial cells where the demand for adenosine triphosphate (ATP) exceeds its production, leading to a reduced ATP/ADP ratio and diminished adenylate energy charge (AEC). This energy deficit impairs growth and essential cellular processes while enhancing glycolysis, oxidative phosphorylation, and reactive oxygen species (ROS) formation [11]. In the context of infectious diseases, bioenergetic stress is critically important because it is intimately linked to the phenomenon of bacterial persistence. Persisters are a subpopulation of dormant, metabolically quiescent bacterial cells that exhibit high tolerance to antibiotics without acquiring genetic resistance [8] [10]. Unlike resistant strains that genetically evade antibiotic action, persisters survive antibiotic exposure by entering a dormant state with reduced metabolic activity and energy consumption, making them insensitive to antibiotics that target active cellular processes [8].
The connection between energy metabolism and antibiotic efficacy is profound. Research has demonstrated that bactericidal antibiotics such as fluoroquinolones, aminoglycosides, and β-lactams induce bioenergetic stress as part of their lethal action, significantly decreasing intracellular ATP, NADH, ATP/ADP ratio, AEC, and NADH/NAD+ ratio [11]. This metabolic disruption creates a hyper-respiratory state that contributes to antibiotic lethality in actively growing cells. However, in persister cells, ATP depletion serves as a protective mechanism that triggers a shift to a dormant-like state, enhancing antibiotic tolerance [8]. Understanding and genetically manipulating these bioenergetic pathways through synthetic biology offers promising strategies to overcome bacterial persistence and address the growing crisis of chronic and relapsing infections that resist conventional antibiotic therapies [10].
Key Pathways and Molecular Mechanisms
Central Energy Metabolism and ATP Homeostasis
The regulation of bacterial bioenergetic status centers on maintaining ATP homeostasis through coordinated activities of central metabolic pathways. The tricarboxylic acid (TCA) cycle serves as the primary hub for energy generation, producing reduced electron carriers that feed into the electron transport chain to drive ATP synthesis through oxidative phosphorylation [16] [11]. Key enzymes in these pathways, including pyruvate dehydrogenase (PDH), alpha-ketoglutarate dehydrogenase (OGDH), and ATP synthase, undergo critical post-translational modifications that regulate their activity. Lipoylation, a highly conserved post-translational modification, is particularly crucial for the proper function of PDH and OGDH complexes, directly linking energy metabolic flux to protein lipoylation status [16].
When ATP consumption exceeds production, cells experience bioenergetic stress characterized by decreased ATP/ADP ratios and AEC. This energy deficit triggers compensatory metabolic responses including enhanced glycolysis and oxidative phosphorylation, leading to increased oxygen consumption and extracellular acidification rates [11]. These metabolic adaptations ultimately result in elevated ROS production, which contributes to oxidative damage of cellular components including DNA, proteins, and lipids. The resulting DNA damage activates stress response pathways that can promote mutagenesis and potentially accelerate resistance development [11].
Signaling Pathways Regulating Persistence and Stress Responses
Table 1: Key Signaling Pathways in Bioenergetic Stress and Bacterial Persistence
| Pathway/System | Key Components | Activation Trigger | Cellular Outcome |
|---|---|---|---|
| Stringent Response | (p)ppGpp, RelA, SpoT | Nutrient limitation, Bioenergetic stress | Growth arrest, Metabolic dormancy, Persister formation |
| Toxin-Antitoxin (TA) Modules | HipA, MazF, RelE | Environmental stress, Antibiotic treatment | Growth inhibition, Persister formation via target inhibition |
| ROS Stress Response | SoxRS, OxyR regulons | Elevated ROS from hyper-metabolism | Oxidative damage protection, DNA repair activation |
| Quorum Sensing (QS) | Autoinducer molecules, Receptor proteins | Population density signals | Biofilm formation, Collective stress response |
| SOS Response | RecA, LexA | DNA damage | DNA repair, Mutagenesis, Potential resistance evolution |
Bacterial cells employ sophisticated signaling networks to sense and respond to bioenergetic challenges. The stringent response, mediated by the alarmones (p)ppGpp, serves as a master regulator during nutrient limitation and energy stress [10] [11]. Under bioenergetic stress conditions, elevated (p)ppGpp levels trigger extensive transcriptional reprogramming that downregulates energy-intensive processes such as ribosome biogenesis, DNA replication, and cell division while upregulating stress resistance mechanisms. This reallocation of cellular resources promotes a dormant, persistent state that enhances survival under adverse conditions, including antibiotic exposure [11].
The toxin-antitoxin (TA) systems constitute another crucial regulatory network connecting bioenergetic status to persistence formation [10]. These modular genetic systems consist of a stable toxin protein that can disrupt essential cellular processes and a labile antitoxin that neutralizes the toxin under normal conditions. Under stress conditions, antitoxins are degraded, allowing toxins to act on their targets. For example, HipA toxin inhibits glutamate metabolism and translation, while MazF cleaves mRNA molecules, both leading to growth arrest and persistence [10]. Multiple TA systems can be activated simultaneously under bioenergetic stress, amplifying the persistence response through synergistic effects on cellular physiology.
Diagram 1: Signaling pathways linking bioenergetic stress to bacterial persistence and resistance evolution. Bioenergetic stress activates multiple signaling systems that collectively promote persister formation and can accelerate resistance development.
Synthetic Biology Approaches to Engineer Bioenergetic Stress
Genetic Tools for Inducing and Controlling Bioenergetic Stress
Synthetic biology provides precise genetic tools to engineer bioenergetic stress for research and therapeutic applications. A well-validated approach involves constructing genetic circuits for inducible ATP hydrolysis. This typically employs constitutive overexpression of Escherichia coli's soluble ATP synthase F1 complex (atpAGD genes), which creates continuous ATP hydrolysis independent of the normal proton motive force-coupled ATP synthesis [11]. This engineered system significantly decreases intracellular ATP/ADP ratios and AEC, mimicking natural bioenergetic stress conditions without the pleiotropic effects of chemical stressors. Similarly, heterologous expression of Streptococcus pneumoniae NADH oxidase (nox) induces constitutive NADH oxidation, decreasing NADH/NAD+ ratios and creating redox stress that indirectly affects energy metabolism [11].
Advanced synthetic biology approaches also leverage lipoylation pathway engineering to modulate energy metabolism across diverse organisms. While bacteria typically possess both de novo and salvage pathways for protein lipoylation, eukaryotes generally rely solely on the de novo pathway [16]. Introducing Escherichia coli-derived lipoate protein ligase A (LplA) from the bacterial salvage pathway into eukaryotic cells (including mammalian, algal, and fungal cells) enhances lipoylation of key metabolic enzymes including PDH and OGDH, leading to improved energy metabolism, mitochondrial function, and overall cell physiology [16]. This PTM-based engineering strategy represents a powerful approach to manipulate central energy metabolism through a single genetic intervention.
Reprogramming Metabolic Flux for Controlled Energy Disruption
Strategic redirection of metabolic flux provides another synthetic biology approach to engineer bioenergetic stress. This can be achieved through promoter engineering to control the expression of key metabolic enzymes, CRISPR-based gene activation or interference (CRISPRa/i) to fine-tune metabolic pathway activity, and synthetic protein degradation tags to precisely control the abundance of metabolic enzymes [33] [34]. For example, downregulating glycolytic enzymes while upregulating TCA cycle and oxidative phosphorylation components can create an imbalance between ATP production and consumption, leading to controlled bioenergetic stress.
Another innovative approach involves engineering biosensor-controlled genetic circuits that automatically induce stress responses upon detecting specific metabolic signals. These circuits can be designed using metabolite-binding transcription factors that regulate expression of ATP-consuming enzymes or metabolic inhibitors when energy charge decreases below a certain threshold [35] [34]. Such self-regulating systems create dynamic feedback loops that allow real-time control of bioenergetic status, enabling more precise studies of how energy fluctuations influence antibiotic efficacy and persistence development.
Experimental Models and Methodologies
Establishing Bioenergetic Stress Models: Protocols and Applications
Table 2: Quantitative Effects of Bioenergetic Stress on Bacterial Physiology and Antibiotic Efficacy
| Parameter Measured | Experimental System | Change vs Control | Functional Significance |
|---|---|---|---|
| ATP/ADP Ratio | E. coli pF1 (ATP hydrolysis) | Significant decrease | Indicates bioenergetic stress state |
| Adenylate Energy Charge | E. coli pF1 (ATP hydrolysis) | Significant decrease | Reduced energy status of cells |
| NADH/NAD+ Ratio | E. coli pNOX (NADH oxidation) | Significant decrease | Indicates redox stress |
| Oxygen Consumption | E. coli pF1 and pNOX | Significant increase | Enhanced respiratory activity |
| Ciprofloxacin Persisters | E. coli pF1 + ciprofloxacin | 63-217x increase | Enhanced antibiotic tolerance |
| Resistance Evolution | E. coli pF1 serial passage | Significantly accelerated | Faster resistance development |
| Intracellular ATP | S. aureus + 10 mM quercetin | 36% decrease | Metabolic stress induction |
Protocol 1: Genetic Engineering of Bioenergetic Stress Using Inducible ATP Hydrolysis System
This protocol establishes a controlled system for inducing bioenergetic stress in Escherichia coli through constitutive ATP hydrolysis:
Plasmid Construction: Clone the atpAGD genes encoding the F1 subunit of ATP synthase into a low-copy number plasmid under a constitutive promoter (e.g., J23100 from the Anderson collection). Include an empty vector with the same backbone as control [11].
Strain Transformation: Introduce the constructed plasmid and empty vector control into E. coli MG1655 or other desired strains using standard heat-shock or electroporation methods. Select transformants on LB agar with appropriate antibiotics.
Validation of Bioenergetic Stress:
- Measure intracellular ATP, ADP, and AMP levels using LC-MS/MS. Calculate ATP/ADP ratio and adenylate energy charge [AEC = (ATP + 0.5ADP)/(ATP + ADP + AMP)] [11].
- Confirm decreased ATP/ADP and AEC in pF1 strains compared to empty vector controls.
- Assess physiological effects by measuring growth kinetics (extended lag phase expected) and metabolic fluxes (increased oxygen consumption rate and extracellular acidification rate) [11].
Antibiotic Persistence Assays: Grow overnight cultures of pF1 and control strains in appropriate medium. Dilute and treat with ciprofloxacin (18 ng/mL, ~1.5x MIC) or other antibiotics for 4-24 hours. Determine surviving cells by plating serial dilutions on drug-free media and counting colonies after 24-hour incubation. Calculate persister fractions as (CFU/mL after treatment)/(CFU/mL before treatment) [11].
Protocol 2: Metabolite-Induced Reprogramming of Bacterial Persisters
This approach utilizes metabolites to reverse the dormant state of persister cells and enhance antibiotic efficacy:
Persister Cell Isolation: Generate persister cells by treating stationary-phase cultures with high concentrations of bactericidal antibiotics (e.g., 10x MIC of ciprofloxacin or ampicillin for 4 hours). Wash cells to remove antibiotics and resuspend in fresh medium [10].
Metabolite Treatment: Prepare metabolite solutions (e.g., 10-100 mM pyruvate, mannitol, or fructose) in appropriate buffers. Add metabolites to persister cell suspensions at final concentrations based on experimental requirements [10].
Metabolic Activation Monitoring:
- Measure intracellular ATP levels using luminometric assays after 1-2 hours of metabolite treatment.
- Assess membrane potential using fluorescent dyes like DiOC₂(3) and flow cytometry.
- Evaluate respiratory activity through oxygen consumption rates using extracellular flux analyzers [10].
Antibiotic Resensitization Assay: Following metabolite pretreatment (1-2 hours), add appropriate antibiotics at determined concentrations. Incubate for additional 4-24 hours and quantify surviving cells by plating and colony counting. Include controls without metabolites and without antibiotics [10].
Diagram 2: Experimental workflow for establishing bioenergetic stress models. The process begins with genetic engineering to create stress systems, followed by validation and application in persistence and resistance studies.
Analytical Methods for Quantifying Bioenergetic Parameters
Accurate measurement of bioenergetic parameters is essential for evaluating engineered stress systems and their effects on bacterial physiology. Metabolite profiling using LC-MS/MS provides comprehensive assessment of energy status by quantifying ATP, ADP, AMP, NAD+, NADH, and key intermediates from central metabolic pathways [11]. Sample preparation involves rapid quenching of metabolism (e.g., cold methanol extraction), followed by analysis using reverse-phase or HILIC chromatography coupled to high-resolution mass spectrometry. This approach enables simultaneous measurement of ATP/ADP ratios, AEC, and NADH/NAD+ ratios, offering a complete picture of cellular energy and redox states.
Metabolic flux analysis provides functional assessment of bioenergetic stress through real-time measurements of respiration and glycolysis. Extracellular flux analyzers enable parallel measurement of oxygen consumption rate (OCR, indicating mitochondrial respiration) and extracellular acidification rate (ECAR, largely representing glycolytic flux) in living cells [11]. For bacterial studies, cultures are seeded in specialized microplates and baseline measurements recorded before and after sequential injection of metabolic inhibitors (e.g., antimycin A for electron transport chain inhibition) or modulators. Bioenergetically stressed cells typically exhibit both enhanced OCR and ECAR compared to controls, reflecting increased energy demand and compensatory metabolic activity.
Additional functional assays include:
- ROS measurement using fluorescent probes like H₂DCFDA followed by flow cytometry or fluorimetry
- Membrane potential assessment with dyes such as DiOC₂(3) and JC-1
- ATP production rates using luciferase-based assays
- Gene expression analysis of stress response pathways via RNA-seq or RT-qPCR
Research Reagents and Experimental Tools
Table 3: Essential Research Reagents for Bioenergetic Stress Studies
| Reagent/Tool | Type | Function/Application | Examples/Sources |
|---|---|---|---|
| pF1 Plasmid System | Genetic construct | Constitutive ATP hydrolysis; induces bioenergetic stress | atpAGD genes in low-copy plasmid [11] |
| pNOX Plasmid System | Genetic construct | Constitutive NADH oxidation; induces redox and energy stress | nox gene from S. pneumoniae [11] |
| LplA Expression System | Genetic tool | Enhances lipoylation of metabolic enzymes; improves energy metabolism | E. coli lipoate protein ligase A [16] |
| Metabolite Standards | Chemical reagents | LC-MS/MS quantification of energy parameters | ATP, ADP, AMP, NAD+, NADH commercial standards |
| Energy Phenotype Assay Kit | Functional assay kit | Simultaneous measurement of OCR and ECAR | Commercial extracellular flux assay kits |
| ROS Detection Probes | Fluorescent dyes | Quantification of reactive oxygen species | H₂DCFDA, CellROX reagents |
| ATP Quantitation Assays | Luminometric kits | Measurement of intracellular ATP levels | Luciferase-based commercial kits |
| CRISPRi/a Systems | Genetic modulation | Targeted gene knockdown/activation for metabolic genes | dCas9-based systems for prokaryotes [34] |
The investigation of bioenergetic stress requires specialized reagents and tools designed to manipulate and measure cellular energy status. Genetic constructs for inducing controlled bioenergetic stress include plasmid systems for constitutive ATP hydrolysis (pF1) and NADH oxidation (pNOX), which can be deployed in various bacterial species with appropriate modification [11]. For eukaryotic systems, mitochondrial-targeted versions of these constructs can be created by adding mitochondrial targeting sequences (e.g., from ornithine transcarbamylase or superoxide dismutase 2) to ensure proper localization [16].
Advanced genome engineering tools enable precise manipulation of energy metabolic pathways. CRISPR-Cas systems, particularly CRISPR interference (CRISPRi) and activation (CRISPRa), allow targeted knockdown or overexpression of genes involved in ATP production, consumption, and regulation [33] [34]. Mobile-CRISPRi systems facilitate functional genomics studies of essential energy metabolism genes across diverse bacterial pathogens, including Pseudomonas aeruginosa, Acinetobacter baumannii, and Vibrio species [34]. These tools enable systematic dissection of how individual metabolic enzymes and pathways contribute to bioenergetic stress and antibiotic persistence.
Analytical platforms for quantifying bioenergetic parameters include extracellular flux analyzers for real-time metabolic phenotyping, LC-MS/MS systems for comprehensive metabolite profiling, and fluorescent imaging systems for spatial assessment of energy status in individual cells or complex microbial communities. Combining these tools with synthetic biology approaches creates a powerful toolkit for understanding and manipulating bioenergetic stress in bacterial pathogens, ultimately leading to new therapeutic strategies against persistent infections.
The synthetic biology approaches to engineer and manipulate bioenergetic stress represent a promising frontier in combating bacterial persistence and antibiotic tolerance. By developing precise genetic tools to control cellular energy status, researchers can systematically investigate the relationship between metabolic state and antibiotic efficacy, identifying vulnerable points in bacterial physiology that can be targeted to overcome treatment failures in chronic and recurrent infections. The experimental models and methodologies outlined provide a framework for advancing this emerging field, enabling both basic research into persistence mechanisms and translational development of novel therapeutic strategies.
Future directions in this field will likely focus on developing more sophisticated genetic circuits that can sense and respond to bioenergetic parameters in real-time, creating dynamic systems that maintain optimal stress levels for resensitizing persisters without promoting resistance evolution [10] [11]. Additionally, applying these approaches to clinical isolates and in vivo infection models will be essential for translating laboratory findings to therapeutic applications. Combining bioenergetic manipulation with conventional antibiotics and other adjuvant approaches offers a multifaceted strategy to address the complex challenge of bacterial persistence, potentially extending the useful lifespan of existing antibiotics and improving outcomes for patients with difficult-to-treat infections. As synthetic biology tools continue to advance, so too will our ability to precisely engineer bacterial metabolism for therapeutic benefit, ultimately strengthening our arsenal against antimicrobial resistance.
The study of bacterial dormancy represents a critical frontier in combating persistent infections, with metabolic flux at its core. Dormant bacterial populations, known as persister cells, contribute significantly to chronic and recurrent infections by entering a state of metabolic quiescence that confers tolerance to conventional antibiotics [8]. This whitepaper examines the central role of energy metabolism reprogramming during dormancy transitions, with particular focus on ATP depletion as a key driver of this phenotypic switch. Understanding these metabolic dynamics provides crucial insights for developing novel therapeutic strategies against persistent bacterial infections.
The fundamental link between ATP depletion and bacterial persistence has emerged as a pivotal area of investigation. Research demonstrates that reduced intracellular ATP levels trigger a shift to a dormant-like state that enhances antibiotic tolerance [8]. This metabolic reprogramming involves complex alterations to metabolic networks that can be quantitatively mapped using advanced metabolomic profiling techniques. Within this framework, this technical guide explores methodologies for analyzing these metabolic transitions, presents key quantitative findings, and discusses implications for drug development targeting persistent bacterial populations.
Metabolic Flux Analysis: Core Methodologies
Mapping metabolic flux during dormancy transitions requires sophisticated analytical and computational approaches that can capture the dynamic rerouting of metabolites through biochemical networks. The table below summarizes the primary techniques employed in this field.
Table 1: Techniques for Determining Metabolic Fluxes
| Technique | Network Scale | Metabolic System | Key Advantages | Major Limitations |
|---|---|---|---|---|
| 13C-MFA (13C-Metabolic Flux Analysis) | Central carbon metabolism | Steady-state | • Independent of reaction kinetics• Considered gold standard for flux determination• High flux precision | • High computational cost• Requires tracer experiments & isotopic measurements• Limited predictive capability |
| FBA (Flux Balance Analysis) | Genome-scale metabolism | Steady-state | • Genome-scale coverage• Low computational cost• Independent of reaction kinetics | • Dependent on biological objective function• Does not incorporate gene regulation data |
| Kinetic Modeling | Small scale to central carbon metabolism | Dynamic | • Mechanistic approach• Captures dynamic behavior• Enables detailed predictions | • High computational cost• Difficult parameterization• Requires extensive experimental data |
| E-Flux | Genome-scale metabolism | Steady-state | • Integrates gene expression data with metabolic models• Predicts metabolic capacity from expression data | • Limited to modeling maximum flux constraints |
These methodologies enable researchers to quantitatively trace the flow of carbon and energy through metabolic networks, providing invaluable information for understanding how bacteria reprogram their metabolism during dormancy transitions [36]. 13C-MFA has emerged as the gold standard for experimental flux determination, utilizing labeled substrates (e.g., 13C-glucose) and analyzing the resulting isotopic patterns in intracellular metabolites to infer metabolic fluxes [36]. This approach depends on mass balance of metabolites and their isotopomers, rather than reaction kinetics, making it particularly valuable for systems where enzymatic parameters are unknown.
Complementarily, constraint-based approaches like FBA leverage genome-scale metabolic models (GEMs) to predict system-level metabolic capabilities. FBA uses a stoichiometric matrix of metabolic reactions and applies constraints based on physical parameters to predict flux distributions that optimize a biological objective, such as biomass production [37]. The E-Flux method extends this framework by incorporating gene expression data to model maximum flux constraints, creating a powerful hybrid approach for predicting metabolic states from transcriptomic measurements [37].
Experimental Protocols for Dormancy Transitions
Investigating Quercetin-Induced Metabolic Stress and Persistence
The following protocol outlines a comprehensive approach for assessing metabolic stress-induced dormancy using quercetin as an inducer of ATP depletion in Staphylococcus aureus [8]:
Bacterial Culture and Treatment:
- Grow S. aureus cultures to mid-exponential phase in appropriate medium.
- Prepare quercetin solutions at varying concentrations (e.g., 1 mM and 10 mM) in suitable solvent.
- Treat experimental cultures with quercetin, maintaining untreated controls.
Intracellular ATP Measurement:
- Harvest bacterial cells by centrifugation at specified time points post-treatment.
- Lyse cells using dedicated ATP extraction reagents.
- Quantify ATP levels using bioluminescence assay based on luciferase-catalyzed light emission.
- Normalize ATP values to cellular protein content or cell count.
Persister Cell Formation Assay:
- Expose quercetin-treated and control cultures to lethal concentrations of antibiotics (e.g., oxacillin, ciprofloxacin, tobramycin).
- Determine surviving bacterial counts by plating on antibiotic-free medium after specified exposure periods.
- Calculate persister frequencies as the ratio of surviving cells to initial population.
Pre-treatment Experimental Design:
- For pre-treatment studies, expose bacterial cultures to quercetin for 1 hour prior to antibiotic challenge.
- Include co-treatment conditions (quercetin and antibiotic administered simultaneously) for comparison.
Single-Cell Metabolic Dynamics in Infected Macrophages
This protocol details the methodology for tracking metabolic dynamics in single infected cells, enabling prediction of bacterial replication outcomes [38]:
Macrophage Infection Model:
- Differentiate human monocyte-derived macrophages (hMDMs) from primary monocytes.
- Infect hMDMs with GFP-expressing Legionella pneumophila at MOI of 10.
- Include control infections with avirulent T4SS mutant (ΔdotA) strains.
Live-Cell Imaging and Metabolic Tracking:
- Plate infected macrophages in 384-well microplates for high-throughput imaging.
- Stain cells with Hoechst (nuclear marker) and Cell Tracker Blue (cytoplasmic marker).
- Acquire time-lapse images at hourly intervals up to 18 hours post-infection using spinning-disk confocal microscopy.
- Monitor metabolic parameters (mitochondrial membrane potential, mROS production) via fluorescent indicators.
Image Analysis and Bacterial Replication Quantification:
- Use BATLI (Backtracking Analysis of Time-Lapse Images) software for retrospective single-cell analysis.
- Track bacterial replication by measuring area occupied by intracellular GFP-expressing bacteria.
- Define replication-positive cells as those showing >15 μm² increase in bacterial area from 0 to 18 hours post-infection.
Machine Learning Model Training:
- Extract temporal metabolic features from single-cell trajectories.
- Train classification models to predict replication outcomes based on early metabolic parameters.
- Validate model accuracy against experimental outcomes.
Quantitative Profiling of Metabolic Flux During Dormancy
The application of metabolic flux analysis to bacterial dormancy has yielded key quantitative insights into the metabolic reprogramming associated with persistence. The following table consolidates significant findings from recent investigations.
Table 2: Quantitative Metabolic Changes During Dormancy Transitions
| Metabolic Parameter | Experimental System | Quantitative Changes | Functional Consequences |
|---|---|---|---|
| Intracellular ATP Levels | S. aureus + Quercetin | • 22% reduction with 1 mM quercetin• 36% reduction with 10 mM quercetin | Dose-dependent metabolic stress inducing dormancy |
| Persister Cell Formation | S. aureus + Quercetin + Antibiotics | • 63-fold increase with oxacillin• 88-fold increase with ciprofloxacin• 217-fold increase with tobramycin | Enhanced antibiotic tolerance across multiple drug classes |
| Bacterial Replication in Single Cells | L. pneumophila in human macrophages | • Replication in 17±8% of infected cells (wild-type)• Replication in 0.4±0.5% of infected cells (ΔdotA mutant) | Host cell heterogeneity determines bacterial replication success |
| Metabolic Prediction Accuracy | Machine learning model of infection | 83% accuracy in predicting bacterial replication at 5 hours post-infection | Early metabolic alterations predict infection outcomes |
These quantitative profiles demonstrate that ATP depletion consistently correlates with enhanced persistence across experimental systems. The dramatic increase in persister cell formation under quercetin-induced metabolic stress—particularly the 217-fold increase observed with tobramycin—highlights the profound impact of energy metabolism on antibiotic efficacy [8]. The single-cell analysis further reveals that only a subset of host cells (approximately 17%) support bacterial replication, emphasizing the critical role of host-pathogen metabolic interactions in determining infection outcomes [38].
Research Reagent Solutions for Metabolic Flux Studies
Table 3: Essential Research Reagents for Metabolic Flux Analysis in Dormancy
| Reagent / Tool | Specific Example | Research Application | Key Function |
|---|---|---|---|
| Metabolic Stress Inducers | Quercetin (1-10 mM) | Inducing metabolic dormancy | Depletes intracellular ATP; triggers bacterial persistence |
| Isotopic Tracers | 13C-glucose, 13C-glutamate | 13C-Metabolic Flux Analysis (13C-MFA) | Enables experimental determination of metabolic fluxes |
| Genome-Scale Model Reconstruction Tools | RAVEN Toolbox, Model SEED, CarveMe | Constraint-based metabolic modeling | Builds simulation-ready metabolic models from genome annotations |
| Live-Cell Imaging Dyes | Hoechst, Cell Tracker Blue, Δψm-sensitive dyes | Single-cell metabolic tracking | Visualizes nuclei, cytoplasm, and mitochondrial parameters in live cells |
| Flux Analysis Algorithms | E-Flux method | Integrating gene expression with metabolic models | Predicts metabolic flux capacity from transcriptomic data |
| Metabolomic Analysis Platforms | GC-MS, LC-HRMS, GNPS Molecular Networking | Comprehensive metabolite profiling | Identifies and quantifies metabolites in complex biological samples |
Metabolic Pathways and Experimental Workflows
Metabolic Flux Regulation Network
Single-Cell Metabolic Tracking Workflow
The mapping of metabolic flux during dormancy transitions reveals ATP depletion as a central regulator of bacterial persistence, creating a paradoxical therapeutic challenge where compounds that initially suppress bacterial growth may inadvertently promote antibiotic tolerance [8]. This understanding enables new approaches for targeting persistent infections, including combination therapies that prevent metabolic dormancy or trigger metabolic activation of dormant cells to enhance antibiotic efficacy.
The integration of single-cell analysis with metabolic flux mapping provides unprecedented resolution for understanding host-pathogen metabolic interactions, offering early predictive markers for infection outcomes [38]. These advances, coupled with genome-scale metabolic modeling and high-resolution metabolomic profiling, create a powerful toolkit for identifying novel drug targets against persistent bacterial populations. Future therapeutic strategies will likely focus on modulating metabolic transitions to prevent dormancy entry or stimulate persistence reversal, ultimately overcoming the limitations of conventional antibiotics against recalcitrant infections.
Bacterial persistence represents a significant challenge in clinical medicine, contributing to chronic and recurrent infections that are difficult to eradicate. Persister cells are growth-arrested, phenotypic variants within isogenic bacterial populations that exhibit multidrug tolerance without genetic resistance [8] [39]. The dormant nature of these cells enables survival against high doses of conventional antibiotics that primarily target actively growing cells. While persistence was once attributed to a uniform state of metabolic dormancy, emerging evidence reveals remarkable metabolic heterogeneity within persister subpopulations that critically influences their survival and resuscitation dynamics [40] [41].
The investigation of metabolic heterogeneity in bacterial persisters has been revolutionized by recent advances in single-cell analytical technologies. These tools have enabled researchers to move beyond bulk population measurements and uncover the significant cell-to-cell variations in metabolite levels and dynamics that exist even under identical environmental conditions [40]. This heterogeneity enables influential population-level functions not possible or measurable at the ensemble scale, representing a fundamental feature of bacterial populations with critical implications for infectious disease treatment and antibiotic development [40].
This technical guide explores the central role of ATP depletion and energy metabolism in shaping persister cell formation, survival, and resuscitation, with a specific focus on how single-cell analysis technologies are revealing unprecedented insights into metabolic heterogeneity. By integrating quantitative data, experimental protocols, and visual schematics, we provide researchers with a comprehensive framework for investigating energy metabolism in persister subpopulations at single-cell resolution.
Metabolic Foundations of Bacterial Persistence
Energy Metabolism and Persister Formation
Adenosine triphosphate (ATP) serves as a fundamental regulator of bacterial persistence, with reduced intracellular ATP levels strongly associated with persister cell formation [8]. ATP depletion triggers a shift to a dormant-like state that enhances antibiotic tolerance by restricting energy-dependent processes required for antibiotic-mediated killing, including transcription, translation, and cell wall synthesis [8]. Single-cell analyses have demonstrated that bacterial populations exhibit significant heterogeneity in ATP levels, forming subpopulations with distinct metabolic activities and antibiotic tolerance profiles [40] [42].
The relationship between ATP depletion and persistence extends beyond a simple binary state. Quantitative measurements reveal that persister cells maintain a spectrum of metabolic activities, with varying degrees of ATP depletion correlating with different levels of antibiotic tolerance [8] [11]. Bioenergetic stress, defined by a decreased ATP/ADP ratio and adenylate energy charge (AEC), creates physiological conditions that potentiate both antimicrobial resistance evolution and persistence [11]. This stress state can be induced by various conditions, including heterologous gene expression, hypoxic stress, oxidative stress, or antibiotic treatment itself [11].
Table 1: Quantitative Measurements of ATP Depletion in Bacterial Persistence
| Experimental Condition | ATP Reduction | Persistence Increase | Organism | Citation |
|---|---|---|---|---|
| Quercetin (1 mM) | 22% | 7-10 fold (varies by antibiotic) | Staphylococcus aureus | [8] |
| Quercetin (10 mM) | 36% | 26-32 fold (varies by antibiotic) | Staphylococcus aureus | [8] |
| Ciprofloxacin (2x MIC) | Significant decrease in ATP/ADP & AEC | Not quantified | Escherichia coli | [11] |
| pF1 bioenergetic stress | Significant decrease in ATP/ADP & AEC | Significant increase across multiple antibiotics | Escherichia coli | [11] |
Mechanisms of Metabolic Heterogeneity
Metabolic heterogeneity in bacterial populations arises from several interconnected biological mechanisms. Molecular noise in gene expression represents a primary source, where stochastic variations in transcription and translation lead to cell-to-cell differences in metabolic enzyme abundance [40]. This variability is particularly pronounced for low-abundance proteins and is influenced by metabolic genes being controlled by noisier promoters compared to essential genes [40]. Positive feedback loops in metabolic regulation can amplify initial stochastic variations, potentially leading to multimodal population distributions where distinct metabolic subpopulations coexist [40].
Asymmetric partitioning of cellular components during cell division represents another key mechanism generating metabolic heterogeneity. Although most small-molecule metabolites exist in high copy numbers with rapid diffusion kinetics, making direct asymmetric distribution unlikely, asymmetric segregation of low-copy-number components can create metabolic differences [40]. This includes uneven distribution of metabolic enzymes, transcription factors regulating metabolic pathways, or subcellular components like inclusion bodies that indirectly affect metabolic activity [40]. These inherited differences may not manifest immediately but can develop over several generations as metabolic networks reach new steady states [40].
Diagram 1: Origins of metabolic heterogeneity in bacterial populations. Three primary mechanisms generate cell-to-cell metabolic variation that influences persister formation: stochastic enzyme expression, positive feedback loops creating bistability, and asymmetric partitioning during division.
Single-Cell Analytical Technologies
Genetically Encoded Biosensors
Genetically encoded metabolite biosensors represent a powerful approach for monitoring metabolic heterogeneity in living bacterial cells by coupling concentrations of specific metabolites to quantitative fluorescent outputs [40]. These sensors enable dynamic tracking of metabolite levels through either flow cytometry or live-cell imaging, with fluorescence-activated cell sorting (FACS) allowing physical separation of subpopulations based on metabolic characteristics [40].
Several design strategies exist for constructing these biosensors. Metabolite-responsive transcription factors can regulate expression of fluorescent reporters or RNA-based fluorescent aptamers, providing transcriptional-level sensing [40]. For faster reporting kinetics, Förster resonance energy transfer (FRET)-based biosensors utilize metabolite-binding induced conformational changes that alter proximity between donor and acceptor fluorescent proteins [40]. RNA aptamer-based systems offer an alternative rapid reporting approach, where metabolite-binding stabilizes adjacent RNA-based fluorescent aptamers [40]. While genetically encoded biosensors provide exceptional temporal resolution in living cells, they are limited to genetically tractable organisms and require careful calibration to ensure quantitative accuracy.
Table 2: Single-Cell Technologies for Metabolic Analysis of Persisters
| Technology | Key Features | Spatial Resolution | Metabolic Coverage | Throughput |
|---|---|---|---|---|
| Genetically Encoded Biosensors | Live-cell monitoring, dynamic tracking, requires genetic modification | Diffraction-limited | Specific metabolites (design-dependent) | High (flow cytometry) |
| NanoSIMS | Elemental and isotopic analysis, subcellular resolution, quantitative | ~50 nm | Limited to elements and stable isotopes | Low |
| SCLIMS (Single-Cell Live Imaging + MS) | Cross-modality, links metabolome to phenotype, untargeted | Single cell | Broad metabolome coverage | Medium |
| BaSSSh-seq (scRNA-seq) | Transcriptional profiling, biofilm applications, assesses regulatory networks | Single cell | Indirect metabolic inference via transcripts | High |
Mass Spectrometry-Based Approaches
Mass spectrometry techniques provide versatile, quantitative metabolite assessment without requiring genetic modification. Nanoscale secondary ion mass spectrometry (NanoSIMS) offers exceptional subcellular resolution (~50 nm) and the ability to measure multiple metabolic species simultaneously, making it particularly valuable for environmental microbiology applications and non-cultivable bacteria [40]. This technology has revealed extensive heterogeneity in processes such as nitrogen fixation and oleic acid accumulation in wastewater sludge bacteria [40].
The recently developed SCLIMS (single-cell live imaging with mass spectrometry) platform represents a cross-modality innovation that integrates single-cell mass spectrometry with live-cell imaging [42]. This approach simultaneously captures metabolomic features and phenotypic characteristics of individual cells, enabling direct correlation between metabolic states and parameters such as oxidative stress levels [42]. The methodology involves incubating cells with fluorescent probes (e.g., DCFDA for oxidative stress), followed by microscopic imaging, single-cell sampling via patch clamp techniques, and subsequent mass spectrometry analysis [42]. This integrated platform maintains cellular viability and metabolism while generating paired datasets of metabolomic and phenotypic information.
Single-Cell RNA Sequencing
Bacterial single-cell RNA sequencing (scRNA-seq) has emerged as a powerful tool for investigating transcriptional heterogeneity in bacterial populations, including persister cells and biofilm communities [43]. The BaSSSh-seq (bacterial scRNA-seq with split-pool barcoding, second strand synthesis, and subtractive hybridization) method represents an advanced implementation optimized for bacterial cells with low metabolic activity characteristic of persistence states [43]. This technique employs split-pool barcoding to label individual cells' transcripts without requiring sophisticated commercial equipment, uses random hexamers for unbiased RNA capture, incorporates second strand synthesis to improve cDNA yield, and implements enzyme-free rRNA depletion to significantly reduce ribosomal RNA contamination [43].
Application of BaSSSh-seq to Staphylococcus aureus biofilms has captured extensive transcriptional heterogeneity and identified distinct subpopulations with varying metabolic activities and stress responses [43]. The method enables researchers to visualize transcriptional regulatory networks across heterogeneous biofilm subpopulations and identify gene sets associated with transitions from planktonic to biofilm growth, providing unprecedented insights into the metabolic adaptations underlying persistence [43].
Experimental Protocols for Single-Cell Metabolic Analysis
SCLIMS Protocol for Cross-Modality Metabolic Analysis
The SCLIMS protocol enables simultaneous assessment of single-cell metabolomes and oxidative states through these key steps [42]:
Cell Culture and Treatment: Culture HEK293T cells (or bacterial cells with adaptation) and induce oxidative stress using 80 μM H₂O₂ in culture medium for 1 hour. Replace with fresh medium and allow 48-hour recovery before analysis.
Oxidative Stress Labeling: Incubate cells with 25 μM DCFDA in buffer for 25 minutes at 37°C. DCFDA is a cell-permeable dye that becomes fluorescent upon oxidation by reactive oxygen species.
Live-Cell Imaging: Image cells using fluorescence microscopy with standard FITC filters (excitation/emission: 495/529 nm). Calculate fluorescent intensity for each cell to quantify oxidative stress levels.
Single-Cell Sampling: Harvest individual cells using patch clamp micropipettes under visual guidance. Transfer each cell directly to mass spectrometry analysis.
Mass Spectrometry Analysis: Perform single-cell metabolomic profiling using high-resolution mass spectrometry (e.g., FTICR-MS) with m/z range 67-1000. Annotate metabolites using HMDB database matching and confirm identities with MS/MS fragmentation.
Data Integration: Create paired datasets linking each cell's metabolomic profile with its corresponding oxidative stress level for cross-modality correlation analysis.
This protocol maintains cellular viability and metabolism throughout the process, with verification experiments confirming that DCFDA incubation does not significantly alter cellular metabolome based on PCA, UMAP, and t-SNE analyses of incubated versus non-incubated cells [42].
Bacterial Persister Isolation and Metabolic Profiling
For targeted metabolic analysis of bacterial persisters, the following protocol enables isolation and characterization [8] [11]:
Persister Induction: Treat mid-exponential phase Staphylococcus aureus cultures (OD₆₀₀ ≈ 0.5) with 10 mM quercetin for 2 hours to induce ATP depletion and persister formation via metabolic stress.
Antibiotic Selection: Add ciprofloxacin (5-10× MIC) to the culture and incubate for 24 hours to eliminate non-persister cells while preserving persisters.
Persister Collection: Centrifuge cultures at 4,000 × g for 10 minutes, wash twice with sterile phosphate-buffered saline (PBS) to remove antibiotics, and resuspend in fresh medium.
ATP Measurement: Lyse persister cells and measure intracellular ATP levels using luciferase-based ATP assay kits according to manufacturer protocols. Normalize ATP values to protein content or cell count.
Metabolic Activity Assessment: Incubate persister cells with 5 μM CFDA-SE (carboxyfluorescein diacetate succinimidyl ester) for 30 minutes at 37°C, followed by flow cytometry analysis to measure esterase activity as a proxy for general metabolic activity.
Resuscitation Monitoring: Dilute persister cells in fresh medium and monitor regrowth through optical density measurements (OD₆₀₀) every 2 hours for 24 hours to assess heterogeneity in resuscitation kinetics.
This protocol enables researchers to isolate persister subpopulations, quantify their metabolic states, and monitor their heterogeneous resuscitation patterns, providing insights into the relationship between metabolic activity and persistence.
Metabolic Regulation of Persister Dynamics
Bioenergetic Stress and Stringent Response
Bioenergetic stress, characterized by decreased ATP/ADP ratios and adenylate energy charge, potentiates persister cell formation through activation of the stringent response [11]. Engineering Escherichia coli for constitutive ATP hydrolysis through overexpression of the F1 ATP synthase complex (atpAGD) or NADH oxidation via heterologous expression of NADH oxidase (nox) creates sustained bioenergetic stress that significantly increases persister fractions across multiple antibiotic classes, including ciprofloxacin, gentamicin, and ampicillin [11]. This enhanced persistence depends on (p)ppGpp signaling, which reprograms cellular metabolism toward energy conservation and growth arrest under nutrient limitation or stress conditions [11].
The relationship between ATP depletion and persistence demonstrates significant context dependence. While moderate ATP depletion promotes persistence through metabolic slowdown and dormancy induction, severe ATP depletion can potentiate antibiotic lethality by disrupting essential maintenance functions [8] [11]. This biphasic relationship creates a complex landscape where the extent and duration of energy deficit determine cellular fate, contributing to the heterogeneous responses observed within persister populations.
Diagram 2: Bioenergetic stress pathway leading to persister formation. ATP and NADH depletion decrease key energy ratios, triggering bioenergetic stress that increases respiration, glycolysis, and ROS production, ultimately activating the stringent response and persister formation.
Metabolic Heterogeneity in Antibiotic Tolerance
Single-cell analyses have revealed that metabolic heterogeneity generates subpopulations with distinct antibiotic tolerance profiles, fundamentally challenging the notion of persistence as a uniform physiological state [40] [42]. Quantitative measurements demonstrate that bacterial populations contain cells with varying levels of metabolic activity that correlate with differential survival under antibiotic exposure [42]. For example, SCLIMS analysis has revealed inverse correlations between cellular ATP levels and oxidative stress, with high-ATP cells showing greater resistance to oxidative challenges but increased susceptibility to antibiotics that target energy-dependent processes [42].
This metabolic heterogeneity creates a bet-hedging strategy that enhances population survival in fluctuating environments [40]. Under stable conditions, metabolically active subpopulations dominate through faster growth, while under stress, dormant subpopulations survive and potentially resuscitate once conditions improve [40]. The dynamic interplay between these subpopulations, mediated by metabolic heterogeneity, ensures population resilience across diverse environmental challenges.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Single-Cell Analysis of Metabolic Heterogeneity
| Reagent/Category | Function/Application | Example Specifics | Experimental Context |
|---|---|---|---|
| DCFDA (Dichlorodihydrofluorescein diacetate) | Fluorescent detection of cellular oxidative stress | 25 μM incubation for 25 min, Ex/Em: 495/529 nm | SCLIMS protocol for cross-modality analysis [42] |
| Quercetin | Induces ATP depletion and metabolic stress for persister formation | 1-10 mM treatment, dose-dependent ATP reduction (22-36%) | Staphylococcus aureus persister induction [8] |
| Ciprofloxacin | Fluoroquinolone antibiotic for persister selection and studies | 5-10× MIC for selection, 16-18 ng/mL (~2× MIC) for E. coli metabolic profiling | Persister isolation & bioenergetic stress studies [11] |
| pF1 Plasmid System | Constitutive ATP hydrolysis to induce bioenergetic stress | E. coli atpAGD genes on low-copy plasmid | Bioenergetic stress research in E. coli [11] |
| pNOX Plasmid System | Constitutive NADH oxidation to induce bioenergetic stress | S. pneumoniae nox gene on low-copy plasmid | Bioenergetic stress research in E. coli [11] |
| CFDA-SE (Carboxyfluorescein diacetate succinimidyl ester) | Measures general metabolic activity via esterase activity | 5 μM incubation for 30 min, flow cytometry analysis | Metabolic activity assessment in persisters [8] |
| BaSSSh-seq Reagents | Bacterial single-cell RNA sequencing with rRNA depletion | Split-pool barcoding, second strand synthesis, subtractive hybridization | Transcriptional heterogeneity in biofilms [43] |
Single-cell analysis technologies have fundamentally transformed our understanding of metabolic heterogeneity in bacterial persister subpopulations, revealing complex relationships between ATP depletion, energy metabolism, and antibiotic tolerance. The integration of genetically encoded biosensors, advanced mass spectrometry platforms, and bacterial single-cell RNA sequencing has enabled researchers to move beyond population-level averages and investigate the metabolic diversity that underlies persistence at unprecedented resolution.
These technical advances have demonstrated that metabolic heterogeneity arises from multiple sources, including stochastic enzyme expression, positive feedback loops, and asymmetric partitioning during cell division. This heterogeneity creates a spectrum of metabolic states within bacterial populations, with varying degrees of ATP depletion and metabolic activity determining antibiotic susceptibility and resuscitation potential. The emerging paradigm recognizes persistence not as a binary state but as a dynamic continuum of metabolic activities that enables bacterial populations to survive fluctuating environmental conditions and antibiotic challenges.
Future research directions will likely focus on developing even more sophisticated multi-omics approaches that simultaneously capture metabolic, transcriptional, and proteomic information from individual persister cells, further elucidating the regulatory networks that control metabolic heterogeneity. Additionally, translating these fundamental insights into therapeutic strategies that specifically target metabolic vulnerabilities in persister subpopulations holds promise for addressing the persistent challenge of chronic and recurrent bacterial infections.
A significant challenge in treating bacterial infections is the presence of bacterial persisters—a sub-population of genetically susceptible, dormant cells that exhibit high levels of antibiotic tolerance. Unlike resistance, which raises the minimum inhibitory concentration (MIC), tolerance enables bacteria to survive antibiotic exposure by reducing the rate of killing, often without altering the MIC [10]. These dormant cells can be found in planktonic populations and within biofilms, and are a major contributor to chronic and relapsing infections such as those caused by Escherichia coli, Pseudomonas aeruginosa, Mycobacterium tuberculosis, and Staphylococcus aureus [10] [12]. The core of the persistence problem lies in the metabolic dormancy of these cells. Since most bactericidal antibiotics target active metabolic processes like cell wall synthesis, protein production, and DNA replication, they are largely ineffective against dormant persisters [10] [44]. This intrinsic link between metabolic activity and antibiotic efficacy has directed research toward strategies that manipulate bacterial metabolism to overcome tolerance.
Central to this metabolic theory of persistence is the role of energy metabolism and ATP depletion. A shallow metabolic state, characterized by reduced metabolic activity and diminished energy consumption, is closely linked to antibiotic tolerance [10]. In Staphylococcus aureus, host-induced oxidative stress triggers ATP depletion, which is associated with the formation of protein aggregates and a deeper state of dormancy, as defined by increased lag times upon resuscitation [45]. This ATP depletion has been experimentally shown to regulate dormancy depth critically, influencing the capacity of persisters to resuscitate once the environmental stress is removed [45] [46]. Therefore, the metabolic state, particularly cellular energy levels, serves as a critical regulator of bacterial persistence and a promising target for therapeutic intervention.
Core Mechanism: Metabolic Reprogramming to Overcome Dormancy
The "wake and kill" or "metabolic activation" strategy aims to bypass the limitations of conventional antibiotics by reactivating key metabolic processes in dormant persisters, thereby re-sensitizing them to antibiotic killing [10]. The pioneering work of Allison et al. (2011) established the proof-of-concept for this approach, demonstrating that specific metabolites could potentiate aminoglycoside antibiotics to eradicate bacterial persisters [47]. The central mechanism involves the reversal of metabolic suppression and the restoration of the proton motive force (PMF).
The Critical Role of Proton Motive Force (PMF)
Aminoglycoside antibiotics require active transport into the bacterial cell, a process dependent on a sufficient PMF [47]. Persister cells, in their metabolically downregulated state, lack this critical PMF, preventing antibiotic uptake. Exogenous metabolites that enter upper glycolysis—such as glucose, mannitol, fructose, and pyruvate—are catabolized to generate NADH. The oxidation of NADH by the electron transport chain drives the generation of PMF [10] [47]. This elevated PMF facilitates aminoglycoside uptake, leading to ribosome binding, mistranslation, and ultimately, bacterial death. This mechanism is potentiated by the fact that persisters appear to be "primed" for metabolite uptake and central metabolism, allowing them to resume respiration and generate PMF without immediately resuming growth [47]. The critical relationship between metabolism and antibiotic efficacy is summarized in the diagram below.
Contrasting Metabolic Inhibition Strategies
It is crucial to distinguish the metabolite-enabled approach from strategies that employ metabolic inhibitors. While metabolite supplementation aims to increase metabolic activity to facilitate antibiotic uptake, certain inhibitors can also be effective by depleting the energy required for persister survival and repair. Thioridazine, a PMF inhibitor, demonstrates strong synergy with ofloxacin, particularly in pre- and co-treatment conditions [48]. In contrast, other metabolic inhibitors like chloramphenicol (translation inhibitor), rifampicin (transcription inhibitor), and arsenate (ATP production inhibitor) are often antagonistic when combined with bactericidal antibiotics like ofloxacin [48]. This suggests that persisters damaged by antibiotics may require specific levels of transcription, translation, and energy to mount repair responses for survival. Thus, the relationship between metabolism and antibiotic efficacy is nuanced; both the stimulation of energy metabolism (for certain drug classes) and its strategic inhibition (for others) can be leveraged to eradicate persisters, depending on the antibiotic's mechanism of action.
Key Experimental Models and Data
The efficacy of metabolite adjuvants has been demonstrated across a range of experimental models, from in vitro assays to animal infection models.
In Vitro and Ex Vivo Models
The foundational experiments involve isolating persisters from stationary-phase cultures or biofilms and treating them with a combination of metabolites and antibiotics. The survival fraction is then quantified using colony-forming unit (CFU) counts or flow cytometry.
Table 1: Key Metabolites in Aminoglycoside Potentiation Against Persisters
| Metabolite | Point of Entry | Potentiation Efficacy (E. coli) | Proposed Reason for Efficacy |
|---|---|---|---|
| Glucose | Upper Glycolysis | ~3-log reduction [47] | Generates NADH via glycolysis, drives ETC and PMF |
| Mannitol | Upper Glycolysis | ~3-log reduction [47] | Generates NADH via glycolysis, drives ETC and PMF |
| Fructose | Upper Glycolysis | ~3-log reduction [47] | Generates NADH via glycolysis, drives ETC and PMF |
| Pyruvate | Lower Glycolysis | ~3-log reduction [47] | Feeds directly into TCA cycle, generates NADH |
| Gluconate | PPP/EDP | Low potentiation [47] | Bypasses upper glycolysis; less efficient NADH generation |
The experimental workflow for validating metabolite adjuvants typically follows a multi-step process, from persister generation to mechanistic investigation, as outlined below.
Biofilm and In Vivo Models
The therapeutic potential of this strategy extends to more complex, clinically relevant models. For instance, the combination of mannitol and gentamicin reduced the viability of E. coli biofilms by over four orders of magnitude, a significantly greater effect than ofloxacin alone [47]. This approach has also shown promise in a mouse model of chronic urinary tract infection, where metabolite-aminoglycoside combinations led to improved bacterial eradication compared to antibiotic treatment alone [47]. These findings highlight the translational potential of metabolite-adjuvant therapy for treating stubborn biofilm-associated and chronic infections.
Detailed Experimental Protocols
To facilitate replication and further research, this section provides detailed methodologies for key experiments in the field.
Protocol 1: Assessing Metabolite Potentiation Against Planktonic Persisters
This protocol is adapted from the seminal study by Allison et al. [47].
Persister Isolation:
- Grow the bacterial strain of interest (e.g., E. coli or S. aureus) to stationary phase (typically 24-48 hours in rich broth like LB).
- Treat the culture with a high concentration of a bacteriostatic antibiotic (e.g., 10x MIC of rifampicin) or a cell-wall active antibiotic (e.g., ampicillin) for 3-6 hours to kill the majority of the growing population.
- Centrifuge the culture, discard the supernatant, and wash the pellet gently with sterile saline or phosphate-buffered saline (PBS) to remove the antibiotic. Resuspend the pellet in a minimal medium (e.g., M9).
Metabolite and Antibiotic Treatment:
- Distribute the persister-enriched suspension into separate tubes.
- Supplement each tube with a filter-sterilized metabolite solution (e.g., 10-20 mM of mannitol, glucose, fructose, or pyruvate). Include a negative control with minimal medium only.
- Add the aminoglycoside antibiotic (e.g., gentamicin at 10-100 µg/mL). Include controls with metabolite only and antibiotic only.
- Incubate the tubes for 2-4 hours under appropriate aerobic conditions with shaking.
Viability Assessment:
- After incubation, serially dilute the samples in PBS.
- Plate the dilutions onto rich agar plates (e.g., LB agar) that do not contain antibiotics or metabolites.
- Incubate the plates for 24-48 hours and count the resulting colonies to determine the CFU/mL.
- Calculation: Compare the log-reduction in CFU/mL between the "antibiotic only" group and the "metabolite + antibiotic" group to quantify potentiation.
Protocol 2: PMF and Aminoglycoside Uptake Measurement
This protocol is critical for confirming the mechanistic basis of metabolite potentiation [47].
Sample Preparation: Prepare persister cells as described in Protocol 1 and pre-incubate them with potentiating (e.g., mannitol) and non-potentiating (e.g., ribose) metabolites for 30 minutes.
Inhibition Control (Optional): To confirm PMF dependence, pre-treat a subset of samples with a proton ionophore like carbonyl cyanide m-chlorophenyl hydrazone (CCCP; e.g., 50 µM for 15 minutes) before adding the metabolite.
Fluorescent Staining:
- For PMF Measurement: Use the fluorescent dye DiOC₂(3). Add the dye to the cell suspension according to the manufacturer's instructions. For bacteria, a concentration of 1-10 µM and incubation for 15-30 minutes is typical. Analyze the cells immediately using a flow cytometer. A shift in fluorescence (red/green ratio) indicates a change in membrane potential, a component of PMF.
- For Aminoglycoside Uptake: Use a fluorescently labeled aminoglycoside, such as Texas Red-conjugated gentamicin (Gent-TR). Add Gent-TR to the cell suspensions and incubate for 5-10 minutes. Wash the cells to remove unbound dye and analyze fluorescence intensity via flow cytometry. Higher fluorescence indicates greater antibiotic uptake.
The Scientist's Toolkit: Research Reagent Solutions
Successful research in this field relies on a set of key reagents and tools, as catalogued below.
Table 2: Essential Research Reagents for Investigating Metabolic Adjuvants
| Reagent Category | Specific Examples | Function in Research |
|---|---|---|
| Potentiating Metabolites | D-Mannitol, D-Glucose, D-Fructose, Sodium Pyruvate [47] | To stimulate upper glycolysis, generate NADH, and restore PMF in persister cells. |
| Key Antibiotics | Gentamicin, Kanamycin, Streptomycin (Aminoglycosides) [10] [47] | The primary lethal agents whose uptake is potentiated by metabolites. |
| PMF Disruptors | Carbonyl cyanide m-chlorophenylhydrazone (CCCP) [47] | A positive control to demonstrate PMF is essential for metabolite-enabled killing. |
| Respiratory Inhibitors | Potassium Cyanide (KCN) [47] | Used to inhibit aerobic respiration and confirm its role in the potentiation mechanism. |
| Fluorescent Probes | DiOC₂(3), Tetramethylrhodamine ethyl ester (TMRE), Texas Red-Gentamicin (Gent-TR) [47] | To quantitatively measure PMF and antibiotic uptake using flow cytometry or microscopy. |
| Genetic Tools | Knockout strains (e.g., NADH dehydrogenase mutants ΔndhΔnuoI, cytochrome oxidase mutants) [47] |
To dissect the specific contributions of metabolic pathways and respiratory chain components. |
The strategy of using metabolite adjuvants to resensitize bacterial persisters represents a paradigm shift in tackling antibiotic tolerance. By targeting the core metabolic vulnerability of dormant cells—their lack of PMF—this approach leverages existing antibiotics in a more effective manner. The evidence from in vitro, biofilm, and initial in vivo models is compelling, demonstrating that metabolites like mannitol and pyruvate can dramatically enhance the killing power of aminoglycosides against a range of pathogens without inducing growth resumption [10] [47].
The clinical translation of this approach, however, faces hurdles. Maintaining effective local concentrations of metabolites in complex infectious niches like biofilms or abscesses remains a challenge [10]. Furthermore, the potential for off-target effects on host cells or the gut microbiota requires careful investigation. Future work should focus on optimizing metabolite delivery, identifying safe and effective synthetic analogs of natural metabolites, and exploring combinations with other anti-persister strategies that target ATP depletion or protein aggregation [45] [46] [48]. As our understanding of the intricate relationship between bacterial metabolism and antibiotic efficacy deepens, the development of therapies that intelligently manipulate this relationship holds great promise for overcoming the persistent problem of chronic and relapsing bacterial infections.
Overcoming Therapeutic Challenges: Strategies to Combat Persistence and Bioenergetic Stress
A significant challenge in combating chronic and relapsing bacterial infections is the presence of bacterial persisters—transiently dormant, metabolically quiescent cells that exhibit high tolerance to antibiotics without genetically acquired resistance [10]. Unlike resistant strains, persisters remain genetically susceptible to antibiotics but evade killing due to their altered physiological state, primarily characterized by profound metabolic dormancy and reduced energy consumption [10] [45]. This tolerance allows bacteria to survive antimicrobial exposure and resume growth once the treatment ceases, leading to recurrent infections [10]. Conventional antibiotic strategies often fail against persisters because many antibiotics target active metabolic processes, rendering them ineffective against dormant cells [10]. The "wake and kill" approach has emerged as a promising therapeutic strategy to overcome this limitation. By reactivating key metabolic processes in dormant persisters through exogenous metabolites, this method re-sensitizes them to conventional antibiotic killing, thereby circumventing the limitations of conventional treatments [10].
Molecular Mechanisms of Bacterial Persistence and Dormancy
Key Pathways in Persister Formation
Bacterial persistence is a multifactorial phenomenon regulated by several interconnected molecular mechanisms that induce a state of metabolic quiescence and stress tolerance:
- Toxin–antitoxin (TA) modules: These genetic elements produce a stable toxin and a labile antitoxin. Under stress conditions, the antitoxin degrades, allowing the toxin to disrupt essential cellular processes such as DNA replication, translation, and cell division, thereby inducing dormancy [10].
- (p)ppGpp-mediated stringent response: This global stress response to nutrient limitation leads to rapid (p)ppGpp accumulation, which downregulates energy-consuming processes (like rRNA and protein synthesis) and promotes survival under adverse conditions [10].
- Reactive oxygen species (ROS)-induced stress responses: Host-derived oxidative stress can cause cellular damage that promotes dormancy. In Staphylococcus aureus, host ROS induces variable dormant states in persisters, associated with decreased translation, energy metabolism, and ATP depletion [45].
- Biofilm formation: Structured bacterial communities embedded in an extracellular matrix exhibit spatial heterogeneity in metabolic activity. Nutrient limitation and poor antibiotic penetration within biofilms create microenvironments that favor dormancy [10].
The Central Role of ATP Depletion and Energy Metabolism
A hallmark of the deeply dormant persister state is a significant reduction in cellular ATP levels [45]. This ATP depletion has far-reaching consequences for cellular viability and antibiotic tolerance:
- Translation Repression: Low ATP levels directly limit protein synthesis, a key energy-consuming process [45].
- Protein Aggregation: ATP depletion promotes the formation of protein aggregates, visible as dark foci in bacterial cells, which further contributes to dormancy depth [45].
- Diminished Proton Motive Force (PMF): Low ATP production reduces PMF, impairing the uptake of aminoglycoside antibiotics and other PMF-dependent drugs [10].
- Heterogeneous Dormancy Depth: Persisters exist along a spectrum of dormancy, from "shallow" to "deep" states, with the latter characterized by more severe ATP depletion and longer lag times for resuscitation [45].
Table 1: Key Characteristics of Bacterial Persisters
| Characteristic | Description | Implication for Antibiotic Efficacy |
|---|---|---|
| Metabolic State | Dormant or metabolically quiescent | Reduced susceptibility to antibiotics targeting active processes |
| Genetic Basis | Genetically susceptible to antibiotics | No inherited resistance to overcome |
| ATP Levels | Significantly depleted [45] | Limited energy for drug uptake and metabolic activation |
| Resuscitation | Heterogeneous lag times for regrowth [45] | Variable timeframes for clinical recurrence |
| Prevalence | Found in both planktonic cells and biofilms [10] | Contributes to diverse infection types |
Metabolic Reactivation: The "Wake and Kill" Strategy
Theoretical Foundation
The fundamental principle underlying metabolic reactivation is the positive correlation between bacterial metabolic rate and the efficacy of bactericidal antibiotics [10]. By providing specific exogenous metabolites that reactivate central metabolic pathways, researchers can reverse the dormant state of persisters, making them vulnerable again to conventional antibiotics. This approach leverages the existing antibiotic arsenal while potentially delaying the development of resistance [10].
Specific Metabolite Interventions
Different classes of metabolites have demonstrated efficacy in reprogramming bacterial metabolism and re-sensitizing persisters to antibiotics:
- Central Carbon Metabolites: Exogenous pyruvate and mannitol have been shown to promote gentamicin uptake and bactericidal activity by restoring the proton motive force (PMF) in Vibrio alginolyticus and Pseudomonas aeruginosa biofilms, respectively [10].
- Amino Acids: L-valine promotes phagocytosis to kill multidrug-resistant bacterial pathogens, while phenylalanine enhances the innate immune response to clear ceftazidime-resistant Vibrio alginolyticus in zebrafish models [10].
- Nucleotides: Exogenous adenosine and/or guanosine enhance tetracycline sensitivity of persister cells [10].
- Fatty Acids: Fatty acid-comprising lysine conjugates have shown efficacy against MRSA by disrupting biofilms with no resistance development [10].
Table 2: Exogenous Metabolites in "Wake and Kill" Approaches
| Metabolite Class | Specific Example(s) | Proposed Mechanism of Action | Target Pathogen/Model |
|---|---|---|---|
| Sugar Alcohols | Mannitol | Enhances antibiotic sensitivity by restoring PMF [10] | Pseudomonas aeruginosa biofilms [10] |
| Central Carbon Metabolites | Pyruvate | Promotes gentamicin uptake via PMF restoration [10] | Vibrio alginolyticus [10] |
| Amino Acids | L-valine, Phenylalanine | Promotes phagocytosis; enhances innate immune response [10] | Multidrug-resistant pathogens; Vibrio alginolyticus in zebrafish [10] |
| Nucleotides | Adenosine, Guanosine | Enhances tetracycline sensitivity [10] | Persister cells [10] |
| Fatty Acid Conjugates | Fatty acid-lysine conjugates | Disrupts biofilms [10] | MRSA [10] |
Experimental Approaches and Research Methodologies
Visualizing ATP Dynamics in Single Cells
Monitoring ATP dynamics at the single-cell level is crucial for understanding metabolic reactivation. The ATeam (Adenosine 5'-Triphosphate indicator based on Epsilon subunit for Analytical Measurements) technology enables real-time monitoring of ATP levels inside individual living cells [49].
ATeam Biosensor Construction and Mechanism
This genetically encoded FRET-based indicator employs the ε subunit of bacterial FoF1-ATP synthase sandwiched between cyan (mseCFP) and yellow (mVenus) fluorescent proteins [49]. The ε subunit undergoes a large conformational change upon ATP binding, bringing the fluorescent proteins closer together and increasing FRET efficiency [49]. Different ATeam variants offer varying ATP affinities, with dissociation constants ranging from 7.4 μM to 3.3 mM, allowing measurement of ATP across physiological concentrations [49].
Research Reagent Solutions
Table 3: Essential Research Reagents for Metabolic Reactivation Studies
| Reagent/Tool | Function/Application | Key Features |
|---|---|---|
| ATeam Biosensors [49] | Real-time monitoring of intracellular ATP dynamics in live cells | • FRET-based• Genetically encoded• Subcellular targeting capabilities• High specificity for ATP |
| Exogenous Metabolites [10] | Reactivation of metabolic pathways in dormant persisters | • Pyruvate (central metabolism)• Mannitol (PMF restoration)• Amino acids (immune modulation)• Nucleotides (antibiotic sensitization) |
| Metabolic Inhibitors [50] | Investigation of ATP synthesis pathways and energy metabolism | • Oligomycin A (OXPHOS inhibitor)• Phloretin (glycolysis inhibitor) |
| FRET Imaging Systems | Detection of ATeam biosensor signals | • Capable of ratiometric imaging• Time-lapse capability• High sensitivity detectors |
Protocol for Evaluating Metabolic Reactivation
A standardized approach for assessing metabolite-mediated reactivation of bacterial persisters:
Experimental Workflow for Wake-and-Kill
Persister Generation: Induce persistence using appropriate methods such as:
- Antibiotic treatment (e.g., high doses of bactericidal antibiotics)
- Nutrient starvation in stationary phase cultures
- Oxidative stress exposure [45]
Metabolic 'Wake' Phase:
- Resuspend purified persisters in fresh medium containing specific metabolites (e.g., 5-10 mM pyruvate, mannitol, or other metabolites)
- Incubate for predetermined timepoints (typically 1-4 hours) to allow metabolic recovery
- Monitor ATP dynamics in real-time using ATeam biosensors [49]
Antibiotic 'Kill' Phase:
- Apply appropriate bactericidal antibiotics at relevant concentrations
- Determine time-kill kinetics over 24-48 hours
- Include controls without metabolites and without antibiotics
Efficacy Assessment:
- Enumeration of colony-forming units (CFUs) at various timepoints
- Flow cytometry with viability staining (e.g., propidium iodide)
- Measurement of resuscitation lag times as an indicator of dormancy depth [45]
Therapeutic Applications and Clinical Translation
Current Research Status
The therapeutic potential of metabolite-adjuvant therapy is being explored across various infection models:
- Chronic Biofilm Infections: Metabolites like mannitol significantly enhance antibiotic sensitivity of persister cells in P. aeruginosa biofilms, potentially addressing difficult-to-treat device-related infections [10].
- Intracellular Pathogens: For intracellular S. aureus persisters, metabolic reactivation strategies must account for the host cell environment, where ROS-induced dormancy presents additional challenges [45].
- Multi-Drug Resistant Pathogens: Fatty acid conjugates and amino acid adjuncts show promise against MRSA and other resistant pathogens by simultaneously disrupting tolerance mechanisms and enhancing immune clearance [10].
Challenges in Clinical Implementation
Despite promising preclinical results, several challenges remain in translating metabolic reactivation approaches to clinical practice:
- Local Concentration Maintenance: Maintaining effective local concentrations of metabolites in complex infectious niches remains problematic [10].
- Timing and Dosing Optimization: The optimal timing and dosing regimens for metabolite-antibiotic combinations need careful determination to maximize efficacy while minimizing potential side effects [10].
- Pathogen-Specific and Microenvironment-Specific Approaches: Therapeutic strategies may need customization based on the specific pathogen, infection site, and host factors that influence bacterial metabolic states [45].
- Formulation and Delivery Challenges: Developing stable formulations that deliver both metabolites and antibiotics to the target site presents technical hurdles [10].
Metabolic reactivation through exogenous metabolites represents a promising "wake and kill" strategy to overcome the significant clinical challenge of bacterial persisters. By targeting the fundamental metabolic dormancy that underlies antibiotic tolerance, this approach leverages existing antibiotics while potentially delaying resistance development. The central role of ATP depletion in maintaining the persistent state provides a key target for intervention, with tools like ATeam biosensors enabling detailed investigation of ATP dynamics at single-cell resolution [49].
Future research directions should focus on optimizing metabolite selection for specific pathogen-infectionsite combinations, developing sophisticated delivery systems for sustained local release, and identifying biomarkers that predict which infections are most likely to respond to metabolic reactivation strategies. As our understanding of bacterial metabolism in host environments deepens, particularly regarding how host-derived stresses like ROS shape bacterial dormancy depth [45], more targeted and effective therapeutic approaches can be developed to address the persistent challenge of recurrent and chronic bacterial infections.
A significant challenge in combating bacterial infections is the phenomenon of bacterial persistence, where a sub-population of bacteria survives antibiotic treatment without acquiring genetic resistance. These bacterial persisters are genetically susceptible, dormant cells that can tolerate antibiotic exposure and lead to relapsing infections [12] [51]. Central to the formation and survival of these persisters is a process of protein aggregation induced by various stresses, including antibiotic treatment [52] [53] [21]. Unlike the historically detrimental view of protein aggregation, recent research reveals that bacteria exploit this process as a protective regulatory mechanism to enter a dormant state by strategically shutting down metabolism [52] [21]. This whitepaper examines the critical role of molecular chaperones in reversing this process, facilitating bacterial recovery from dormancy by disentangling protein aggregates, thereby restoring metabolic activity. This knowledge is framed within the crucial context of ATP depletion and energy metabolism, which governs both entry into and exit from the persistent state [8] [46].
The Link Between Energy Metabolism, Protein Aggregation, and Bacterial Dormancy
ATP Depletion as the Trigger for Dormancy
Bacterial entry into dormancy is intimately linked to metabolic downregulation, particularly a sharp decrease in intracellular ATP levels. This energy crisis can be triggered by external stressors, including antibiotic exposure or treatment with bioactive compounds like quercetin. Studies on Staphylococcus aureus have demonstrated that quercetin treatment induces a dose-dependent depletion of intracellular ATP, reducing levels by 22% and 36% at concentrations of 1 mM and 10 mM, respectively [8]. This ATP reduction correlates directly with a transition to a dormant, antibiotic-tolerant state.
Protein Aggregation as a Functional Response
Rather than being a purely pathological process, protein aggregation serves as a functional mechanism for implementing metabolic shutdown. Under stress, specific metabolic proteins undergo liquid-liquid phase separation (LLPS), forming dense condensates that transform into gel-like droplets and eventually solidify [52] [21]. This process effectively sequesters and inactivates core metabolic enzymes, facilitating a coordinated reduction in cellular energy production and consumption. The aggregation process thus creates a protective hibernating state where essential proteins are preserved in an inactive form, shielded from degradation or irreversible damage until favorable conditions return [52].
Table 1: Quantitative Relationship Between ATP Depletion and Persister Formation
| Inducing Factor | ATP Reduction | Impact on Persister Formation | Experimental Organism |
|---|---|---|---|
| Quercetin (1 mM) | 22% | 10-fold increase with oxacillin (pre-treatment) | Staphylococcus aureus [8] |
| Quercetin (10 mM) | 36% | 32-fold increase with oxacillin (pre-treatment) | Staphylococcus aureus [8] |
| Quercetin + Tobramycin | Not specified | 217-fold increase in persisters | Staphylococcus aureus [8] |
| Metabolic Stress | Not specified | Induction of deep dormancy states | Escherichia coli [46] |
Molecular Mechanisms of Chaperone-Mediated Aggregate Disassembly and Reactivation
The reversal of protein aggregation is an active process orchestrated by a network of molecular chaperones, with the DnaK-ClpB bi-chaperone system playing a pivotal role in extracting and refolding aggregated proteins.
The DnaK-ClpB Reactivation Pathway
The recovery from dormancy is an energy-dependent process mediated by a coordinated chaperone system. The DnaK-ClpB system operates through a sophisticated mechanism to disaggregate proteins:
- Recognition and Targeting: DnaK (Hsp70) first recognizes and binds to hydrophobic patches exposed on the surface of protein aggregates [53].
- Collaborative Disaggregation: ClpB (Hsp104), a AAA+ ATPase, is recruited to the aggregate through interactions with DnaK. Using the energy from ATP hydrolysis, ClpB undergoes conformational changes that mechanically pull individual polypeptide chains through its central pore, disentangling them from the aggregate [53] [46].
- Refolding and Reactivation: Once extracted, the partially unfolded polypeptides are transferred to DnaK and its co-chaperones for folding into native conformations, thereby restoring protein function and restarting cellular metabolism [53] [21].
This chaperone-mediated disaggregation is critically dependent on ATP availability, creating a direct link between cellular energy status and the capacity to exit dormancy [46]. When ATP levels are restored, the chaperone system becomes activated, initiating the recovery process.
Diagram 1: The Chaperone-Mediated Protein Disaggregation Pathway. This process illustrates the sequential steps by which the DnaK-ClpB system disentangles aggregated proteins, facilitating bacterial exit from dormancy.
Regulatory Dynamics of the Chaperone Network
The activity of the chaperone network is precisely regulated in response to cellular stress. Under normal conditions, expression of chaperones is maintained at basal levels. However, proteostasis disruption and accumulation of misfolded proteins trigger stress response pathways that upregulate chaperone synthesis [53]. This creates a positive feedback loop where increased chaperone production enhances the cell's capacity to resolve protein aggregates once ATP levels are restored. The ATP-dependent nature of both DnaK and ClpB means that recovery is intrinsically linked to the restoration of energy metabolism, ensuring that bacteria only exit dormancy when conditions favorable for growth return [46].
Experimental Approaches for Studying Protein Aggregation and Persister Recovery
Methodologies for Inducing and Quantifying Protein Aggregation
Research into persister cell biology requires specialized methodologies for inducing dormancy and quantifying protein aggregation:
- Inducing Dormancy and Aggregation: Common methods include antibiotic exposure (e.g., fluoroquinolones, β-lactams), metabolic inhibitors like quercetin [8], and nutrient starvation. These treatments trigger the stringent response and toxin-antitoxin module activation, leading to metabolic shutdown and protein aggregation [12] [51].
- Detecting and Quantifying Aggregates: Advanced techniques include:
- Fluorescence-based assays using dyes like PROTEOSTAT or Thioflavin T that exhibit enhanced fluorescence upon binding to protein aggregates [54].
- High-throughput dynamic light scattering (DLS) for measuring particle size distribution and aggregation state in solution [55].
- Differential scanning fluorimetry (DSF) to monitor protein stability and aggregation propensity under different conditions [55].
- Flow cytometry combined with fluorescent dyes for single-cell analysis of aggregation in bacterial populations [54].
Table 2: Key Experimental Methods for Protein Aggregation Analysis
| Method | Principle | Applications in Persister Research | Throughput Capacity |
|---|---|---|---|
| PROTEOSTAT Assay | Fluorescent dye binding to aggregates | Quantification of aggregation extent in persisters; screening anti-aggregation compounds | High (96/384-well format) [54] |
| Dynamic Light Scattering (DLS) | Measures hydrodynamic radius of particles | Monitoring aggregate size distribution over time; assessing sample monodispersity | Medium to High (plate-based formats) [55] |
| Differential Scanning Fluorimetry (DSF) | Detects thermal denaturation of proteins | Identifying conditions that stabilize proteins against aggregation | High (96-well format) [55] |
| Flow Cytometry | Single-particle analysis using scattering & fluorescence | Characterizing heterogeneity in bacterial populations; distinguishing aggregating cells | Medium [54] |
Protocols for Analyzing Chaperone-Mediated Recovery
To study the recovery process, researchers employ specific protocols focusing on chaperone function and protein refolding:
- Experimental Workflow for Assessing Recovery:
- Persister Induction: Treat bacterial cultures with bactericidal antibiotics (e.g., ciprofloxacin, tobramycin) at 5-10× MIC for 3-6 hours to eliminate growing cells and enrich for persisters [8] [12].
- Aggregate Isolation: Separate protein aggregates from soluble fractions via differential centrifugation or filter-based methods.
- Disaggregation Assay: Incubate aggregates with purified DnaK-ClpB chaperone system in ATP-containing buffer at 37°C.
- Activity Measurement: Monitor restoration of enzymatic activity in recovered proteins or use bacterial regrowth curves to quantify resuscitation [46] [21].
Diagram 2: Experimental Workflow for Studying Persister Formation and Recovery. This flowchart outlines the key stages in investigating the complete cycle of bacterial dormancy entry through protein aggregation and chaperone-mediated recovery.
Research Reagent Solutions for Investigating Persister Biology
Table 3: Essential Research Reagents for Protein Aggregation and Chaperone Studies
| Reagent/Assay | Function/Application | Key Features | Research Utility |
|---|---|---|---|
| PROTEOSTAT Protein Aggregation Assay | Detection and quantification of protein aggregates | High-throughput compatible; sensitive to sub-micromolar aggregates; works across pH 4-10 [54] | Screening conditions that modulate aggregation; monitoring disaggregation kinetics |
| DnaK & ClpB Purification Kits | Isolation of chaperone proteins for in vitro studies | Various commercial systems for recombinant protein expression and purification | Enabling mechanistic studies of disaggregation; reconstituting reactivation pathways |
| ATP Determination Kits | Quantifying intracellular ATP levels | Luminescence-based; highly sensitive | Correlating ATP levels with persistence depth and recovery capacity |
| Quercetin | Inducer of metabolic stress and ATP depletion | Natural flavonoid; dose-dependent ATP reduction (22-36%) [8] | Experimental persister induction; studying metabolic regulation of dormancy |
| HipA7 Mutant Strains | High-persistence bacterial strains | Mutation in toxin-antitoxin system; produces ~1% persisters [12] | Genetic model for studying persistence mechanisms |
Implications for Therapeutic Development and Concluding Perspectives
The intricate relationship between protein aggregation, chaperone-mediated reactivation, and bacterial persistence presents both challenges and opportunities for therapeutic development. The understanding that protein aggregation serves a functional role in bacterial dormancy represents a paradigm shift in our comprehension of bacterial stress adaptation [52] [21]. This mechanism, conserved across various bacterial species including Escherichia coli and Staphylococcus aureus, offers potential novel therapeutic targets for combating persistent infections.
Future therapeutic strategies may focus on inhibiting the chaperone systems responsible for protein disaggregation, particularly the DnaK-ClpB complex, to prevent resuscitation of dormant persisters [53] [46]. Alternatively, approaches that promote irreversible protein aggregation could lock bacteria in a permanent dormant state, effectively achieving microbial stasis. The development of anti-aggregation compounds that disrupt the initial formation of protective condensates represents another promising avenue [56] [21]. However, these approaches must account for the dynamic and heterogeneous nature of persister populations, which exhibit a continuum of dormancy depths regulated by the maturation state of protein aggregates [46] [21].
As research continues to unravel the complex interplay between energy metabolism, protein aggregation, and chaperone activity, new opportunities will emerge for designing combination therapies that specifically target the persistent state. Integrating these approaches with conventional antibiotics holds promise for achieving more complete bacterial eradication and addressing the growing crisis of treatment-resistant recurrent infections.
The eradication of dormant, antibiotic-tolerant bacterial cells represents a critical challenge in managing persistent infections. This technical guide examines the central role of bacterial energy metabolism, particularly ATP depletion, in regulating dormancy depth and subsequent resuscitation capacity. Emerging strategies that strategically combine antimicrobial agents based on their mechanisms of action and timing show promising potential to overcome this therapeutic hurdle. By understanding the physiological state of dormant cells and exploiting their metabolic vulnerabilities, researchers can design sophisticated treatment regimens that prevent regrowth and relapse. The following sections provide a comprehensive analysis of dormancy mechanisms, current therapeutic approaches, and detailed experimental methodologies for evaluating novel combination therapies, with specific focus on ATP homeostasis as a key regulator of bacterial persistence.
The Dormancy Phenomenon and Its Link to Energy Metabolism
Defining Bacterial Dormancy and Persistence
Bacterial dormancy describes a reversible state of low metabolic activity in which bacteria can survive for extended periods without division [45]. Within isogenic populations, a small subpopulation of persister cells can exhibit remarkable tolerance to antibiotic challenges, not through genetic resistance mechanisms, but via phenotypic heterogeneity that enables survival under stress [57]. These antibiotic-tolerant cells are often slow-growing or non-growing, rendering them recalcitrant to conventional antibiotics that primarily target active cellular processes [58] [59]. The dormancy state exists on a spectrum, with cells exhibiting varying dormancy depths that correlate with their resuscitation potential [45] [46].
ATP Depletion as a Central Regulator of Dormancy Depth
Recent research has established a crucial relationship between cellular ATP levels and the depth of bacterial dormancy. ATP depletion promotes protein aggregation, leading to increased lag time for cell resuscitation, which directly correlates with bacterial dormancy depth [45] [46]. In Staphylococcus aureus, host-cell-induced oxidative stress triggers ATP depletion, which is associated with the formation of visible dark foci resembling protein aggregates and recruitment of the DnaK-ClpB chaperone system involved in protein aggregate clearance [45]. This energy crisis creates a continuum of dormancy states, from shallow persisters that readily resuscitate to deep persisters with prolonged lag times, and in extreme cases, transitioning to a viable but non-culturable (VBNC) state [45]. The central role of ATP extends beyond energy currency to a critical regulator of cellular integrity in dormant cells.
Table 1: Bacterial Dormancy States and Characteristics
| Dormancy State | Metabolic Activity | ATP Level | Resuscitation Potential | Antibiotic Tolerance |
|---|---|---|---|---|
| Shallow Persister | Moderately reduced | Moderate | Short lag time | High |
| Deep Persister | Significantly reduced | Low | Long lag time | Very High |
| VBNC State | Minimal/Undetectable | Very Low | Requires specific stimuli | Extreme |
Therapeutic Strategies Targeting Dormant Cells
Membrane-Active Agents and Energy Metabolism Disruptors
Conventional antibiotics that target biosynthetic processes in growing cells are notoriously ineffective against dormant bacteria, creating an urgent need for antimicrobials that can treat infections containing non-growing cells [58]. Membrane-active agents and compounds that disrupt energy metabolism represent promising approaches:
Membrane-Targeting Antimicrobials: Agents such as daptomycin and telavancin disrupt the bacterial membrane bilayer, compromising membrane potential and integrity even in non-growing cells [58]. These lipophilic compounds interact directly with the membrane, causing permeabilization and depolarization that proves lethal to dormant bacteria.
Energy Metabolism Inhibitors: The diarylquinoline TMC207 (bedaquiline) targets membrane-bound ATP synthase in Mycobacterium tuberculosis, disrupting energy production essential for maintaining viability even in the absence of growth [58]. This approach exploits the fundamental requirement for cellular energy and redox homeostasis in all living cells, including quiescent bacteria.
Innovative Phage-Based Approaches
Bacteriophages have emerged as promising tools for targeting dormant bacterial populations. While most well-studied phages enter a hibernation state in dormant hosts, recent discoveries have identified phages capable of direct replication on dormant cells:
Phage Paride: This newly isolated Pseudomonas aeruginosa jumbo phage (287,267 bp genome) uniquely replicates on and lyses deep-dormant host cells, achieving >99% reduction in bacterial population [59]. Unlike conventional phages that remain dormant in non-growing bacteria, Paride directly replicates in deep-stationary phase cultures.
Phage-Antibiotic Synergy: The combination of Paride with the β-lactam meropenem enables complete eradication of deep-dormant cultures in vitro and significantly reduces bacterial loads in murine tissue cage infection models [59]. This synergistic approach represents a paradigm shift in targeting persistent infections.
Strategic Combination Therapy Timing
The sequence and timing of combination therapy administration critically determine treatment efficacy against dormant populations:
Concurrent Administration: Simultaneous application of phage Paride and meropenem demonstrates direct synergy, possibly due to phage-induced compromise of cellular integrity enhancing antibiotic activity [59].
Metabolic Priming: Approaches that force dormant cells to transiently increase metabolic activity before applying conventional antibiotics can reverse tolerance mechanisms [58].
Table 2: Combination Therapies Against Dormant Bacteria
| Therapeutic Combination | Mechanism of Action | Target Bacteria | Experimental Model | Efficacy |
|---|---|---|---|---|
| Phage Paride + Meropenem | Phage lytic replication combined with cell wall synthesis inhibition | Pseudomonas aeruginosa | In vitro culture & murine tissue cage infection | Sterilization of deep-dormant cultures in vitro; significant reduction in vivo |
| Membrane-Active Agents + Standard Antibiotics | Membrane disruption enhances uptake of secondary antibiotic | Gram-positive bacteria (e.g., Staphylococcus aureus) | In vitro biofilm models | Enhanced killing of stationary phase and biofilm cells |
| ATP Synthase Inhibitors + Respiratory Inhibitors | Synergistic disruption of energy metabolism | Mycobacterium tuberculosis | In vitro and intracellular models | Improved killing of dormant populations in granuloma models |
Experimental Protocols for Evaluating Anti-Persister Therapies
Monitoring Dormant Microorganisms via Revived ASV Monitoring (RAM)
Accurate monitoring of dormant microorganisms is essential for evaluating therapy efficacy. The Revived Amplicon Sequence Variants (ASV) Monitoring (RAM) method provides a robust approach for identifying dormant microorganisms in experimental systems:
System Setup: Establish a closed experimental system using a homogeneous medium such as Pao cai soup (Chinese fermented vegetables) to exclude exogenous microorganisms [60].
Sampling Regimen: Collect samples at multiple timepoints over an extended period (e.g., 26 timepoints over 60 days) to capture dynamic presence-absence-presence patterns characteristic of dormant microorganisms [60].
Sequencing Protocol:
- Extract total DNA using kits such as the E.Z.N.A. soil kit (Omega Bio-Tek)
- Amplify the bacterial V3–V4 variable region using 338F/806R primers
- Perform sequence amplification with NovaSeq 6000 PE250 platform
- Analyze ASVs that exhibit presence-disappearance-presence patterns as dormant taxa [60]
Data Analysis: Compare results with gene function prediction (GFP) methods to validate the identification of dormant taxa, with RAM demonstrating superior detection of 27,415 ASVs and 616 genera compared to 5,045 ASVs and 270 genera with GFP [60].
Quantifying Persister Fractions and Dormancy Depth
Precise quantification of persister cells requires standardized methodologies to enable cross-study comparisons:
Two-State Model Application:
- Model cells as existing in either normal (N) or persister (P) states
- Normal cells die at rate μ and switch to persister state at rate α during antibiotic treatment
- Persister cells do not die or grow, switching to normal state at rate β [57]
Dormancy Depth Assessment:
ATP Level Determination:
- Culture bacteria in defined media under conditions that induce dormancy
- Collect samples at multiple growth phases (exponential, early stationary, deep stationary)
- Prepare bacterial extracts using ice-cold perchloric acid extraction method
- Neutralize extracts with KOH/KHCO3 solution
- Determine ATP levels using luciferase-based assays (e.g., BacTiter-Glo Microbial Cell Viability Assay) [61]
Host Cell Models for Intracellular Persistence Studies
For pathogens that establish intracellular infections, such as Staphylococcus aureus, specialized host cell models are required:
Macrophage Infection Models:
- Utilize J774 macrophages (low oxidative stress) and human primary macrophages (high oxidative stress) [45]
- Modulate ROS production using LPS and IFN-γ stimulation (enhancement) or butylated hydroxyanisole (inhibition)
- Infect cells with GFP-expressing bacteria and monitor using flow cytometry
Single-Cell Analysis:
- Track bacterial division via GFP signal dilution during cell division
- Use propidium iodide exclusion to identify viable bacteria
- Correlate host cell ROS production with bacterial dormancy depth [45]
Research Reagent Solutions
Table 3: Essential Research Reagents for Dormancy Studies
| Reagent/Cell Line | Specific Example | Application | Key Function |
|---|---|---|---|
| Bacterial Strains | E. coli K12 MG1655; P. aeruginosa clinical isolates; S. aureus GFP-expressing strains | General persistence studies; intracellular dormancy | Model organisms with characterized persistence behavior |
| Cell Lines | J774 macrophages; Human primary macrophages | Intracellular persistence models | Host cells with varying oxidative stress capabilities |
| Detection Kits | BacTiter-Glo Microbial Cell Viability Assay; E.Z.N.A. soil DNA kit | ATP measurement; DNA extraction | Luciferase-based ATP detection; high-quality DNA extraction for sequencing |
| Culture Media | Defined minimal media (M9); Luria Bertani (LB) broth | Dormancy induction | Reproducible culture conditions for persistence studies |
| Antibiotics | Meropenem; Oxacillin; Ciprofloxacin | Persister selection and challenge | Antibiotics for selecting tolerant subpopulations |
Signaling Pathways and Conceptual Workflows
Host-Induced Dormancy Pathway
Diagram 1: Host-induced dormancy pathway in S. aureus.
Combination Therapy Evaluation Workflow
Diagram 2: Combination therapy evaluation workflow.
Future Research Directions
The development of effective combination therapies against dormant bacterial cells requires addressing several critical research areas. First, comprehensive profiling of ATP dynamics across bacterial species and dormancy states will establish fundamental thresholds governing persistence and resuscitation. Second, systematic screening of phage libraries may identify additional viral candidates capable of infecting dormant cells, expanding our therapeutic arsenal. Third, clinical validation of timing sequences for combination therapies is essential to translate laboratory findings into treatment protocols. Finally, advanced imaging technologies to visualize real-time protein aggregation and membrane integrity in dormant cells will provide crucial insights into mechanisms of action for novel therapeutics. Research prioritizing these areas will significantly advance our capacity to combat persistent bacterial infections.
Bioenergetic stress, characterized by a disruption of cellular energy homeostasis, is emerging as a pivotal mechanism driving the evolution of antimicrobial resistance and the formation of persistent bacterial infections. This technical review examines the intricate relationship between adenosine triphosphate (ATP) depletion, reactive oxygen species (ROS) generation, and DNA damage repair in promoting bacterial survival under antibiotic pressure. We synthesize recent findings demonstrating that bioenergetic stress potentiates resistance evolution through ROS-mediated mutagenesis and enhances persistence via the stringent response. The article provides a comprehensive framework for addressing this challenge, detailing experimental protocols for quantifying bioenergetic parameters, and proposes novel therapeutic strategies combining ROS-management with DNA repair inhibition to overcome treatment-resistant infections.
The global health crisis of antimicrobial resistance (AMR) necessitates a paradigm shift in our understanding of how bacteria survive antibiotic treatment. Beyond genetic resistance mechanisms, a crucial yet underappreciated facet is the role of bacterial metabolic states, particularly those involving energy crisis. Bioenergetic stress is defined as a cellular state where ATP consumption chronically exceeds ATP production, leading to a reduced adenylate energy charge (AEC) and altered redox ratios (NADH/NAD⁺) [11]. This energy deficit triggers compensatory metabolic rewiring that inadvertently accelerates the development of both heritable resistance and non-heritable tolerance.
Within the context of dormant bacteria research, ATP depletion is not merely a consequence of antibiotic action but an active driver of bacterial survival strategies. Counterintuitively, a moderate energy deficit can activate hypermetabolic responses that increase mutagenesis, while a severe deficit can induce a deep dormant state that enhances tolerance [11] [62]. This review delineates the mechanisms by which bioenergetic stress-induced ROS signaling and DNA damage repair interact to foster resistance, and outlines targeted experimental and therapeutic approaches to disrupt this cycle.
Core Mechanisms: Linking Energy Depletion to Resistance and Persistence
The Dual Role of ROS in Stress and Mutagenesis
Reactive oxygen species (ROS) serve as critical signaling molecules and executioners in antibiotic-mediated cell death. However, under conditions of bioenergetic stress, their role becomes complex and paradoxical.
- Origin and Amplification: Antibiotic treatment, particularly with fluoroquinolones, can hyper-stimulate cellular respiration in an attempt to replenish ATP pools. This increased electron flow through the electron transport chain elevates the leak of superoxide anions (O₂⁻) and hydrogen peroxide (H₂O₂) [11] [63]. The constitutive hydrolysis of ATP itself directly increases respiration and ROS formation [11].
- DNA Damage and Mutagenic Repair: The accumulated ROS inflict extensive cellular damage, most critically to the bacterial genome. ROS oxidize nucleotide pools and cause single- and double-strand DNA breaks [11] [63]. This oxidative damage activates stress-induced mutagenesis pathways—specifically mutagenic break repair and transcription-coupled repair—which are inherently error-prone. While aiming to repair the damage, these systems introduce mutations that can confer genetic resistance if they occur in genes related to antibiotic target sites or efflux [11].
Table 1: Quantitative Impact of Bioenergetic Stress on Resistance and Persistence
| Parameter | Experimental System | Change vs. Control | Citation |
|---|---|---|---|
| Ciprofloxacin Resistance Evolution | E. coli with constitutive ATP hydrolysis (pF1) | Significantly accelerated (Area Under Curve increased) | [11] |
| Persister Cell Fraction (Ciprofloxacin) | E. coli with constitutive ATP hydrolysis (pF1) | ≈10-fold increase in survival | [11] [62] |
| Intracellular ATP Level | S. aureus treated with 10 mM Quercetin | 36% decrease | [8] |
| Persister Cell Fraction (Tobramycin) | S. aureus with Quercetin co-treatment | 217-fold increase | [8] |
The Stringent Response and Persistence
Concurrent with ROS-mediated mutagenesis, bioenergetic stress potentiates antibiotic persistence. Persisters are a transiently dormant, antibiotic-tolerant subpopulation that can resuscitate after treatment, facilitating relapse and subsequent resistance evolution [8] [45]. The key mechanistic link is the stringent response [11].
ATP depletion and the resulting decrease in the ATP/ADP ratio are sensed as a starvation signal. This triggers the stringent response, a global regulatory program mediated by the alarmone (p)ppGpp. This response dramatically reprograms cellular physiology, shutting down energy-intensive processes like ribosome biogenesis and cell division, and promoting a dormant, stress-resistant state [11]. This metabolic quiescence protects bacteria from antibiotics that primarily target active cellular processes. The depth of this dormancy, and thus the level of tolerance, is heterogeneous and influenced by the extent of ATP depletion and host-derived stresses like oxidative bursts from macrophages [45].
The following diagram illustrates the core signaling pathway from bioenergetic stress to resistance and persistence outcomes.
Experimental Protocols for Investigating Bioenergetic Stress
Inducing and Validating Bioenergetic Stress
A synthetic biology approach provides a direct method to induce and study bioenergetic stress without the confounding pleiotropic effects of antibiotics.
- Genetic Engineering of ATP/NADH Consumption:
- Plasmid System: Utilize a low-copy plasmid system for constitutive expression.
- ATP Stress: Constitutively express the soluble F1 complex of the E. coli ATP synthase (genes atpAGD). This creates a "short circuit," leading to continuous ATP hydrolysis [11] [62].
- NADH Stress: Constitutively express a heterologous NADH oxidase (e.g., from Streptococcus pneumoniae). This enzyme continuously oxidizes NADH to NAD⁺, disrupting the NADH/NAD⁺ ratio and redox balance [11].
- Controls: Always include an empty vector control (pEmpty) in parallel.
- Validation via Metabolomics:
- Sample Preparation: Harvest exponential-phase cells and perform rapid quenching of metabolism (e.g., cold methanol).
- LC-MS/MS Analysis: Quantify intracellular concentrations of ATP, ADP, AMP, NAD⁺, and NADH.
- Key Calculations:
- ATP/ADP Ratio: A key indicator of energy status.
- Adenylate Energy Charge (AEC): A more comprehensive measure calculated as ( [ATP] + 0.5[ADP] ) / ( [ATP] + [ADP] + [AMP] ). A significant decrease validates bioenergetic stress [11].
- NADH/NAD⁺ Ratio: An indicator of redox stress.
Measuring Functional Outcomes: Resistance and Persistence
- Laboratory Evolution of Resistance:
- Protocol: Subject engineered (pF1, pNOX) and control (pEmpty) strains to serial passage in liquid media with progressively increasing concentrations of an antibiotic (e.g., ciprofloxacin). The concentration is typically doubled every 2-3 cycles once robust growth is observed.
- Data Collection: Monitor the fold-change in Minimum Inhibitory Concentration (MIC) over passage cycles. The Area Under the Curve (AUC) of the MIC increase and the number of cycles to reach a pre-defined resistance threshold (e.g., 4x MIC) are key metrics for comparing the rate of resistance evolution [11].
- Time-Kill Assays for Persistence:
- Protocol: Treat mid-exponential phase cultures with a high concentration of a bactericidal antibiotic (e.g., ~5-10x MIC of ciprofloxacin, gentamicin, or ampicillin). Maintain the culture under these conditions for 24 hours.
- Sampling and Enumeration: Take samples at predetermined timepoints (e.g., 0, 2, 4, 6, 24h). Serially dilute and plate on drug-free agar to enumerate surviving Colony Forming Units (CFUs).
- Analysis: Plot the log₁₀(CFU/mL) over time. A biphasic kill curve, with a rapid initial kill followed by a plateau of surviving cells, indicates persister formation. The persister fraction is calculated as the number of CFUs at 24h divided by the initial CFU count [11] [8].
Table 2: Key Reagent Solutions for Bioenergetic Stress Research
| Reagent / Tool | Function / Mechanism | Example Application |
|---|---|---|
| pF1 Plasmid (atpAGD) | Constitutive ATP hydrolysis; induces bioenergetic stress by lowering ATP/ADP and AEC. | Studying the direct effects of ATP depletion on antibiotic efficacy and resistance evolution. [11] |
| pNOX Plasmid (nox) | Constitutive NADH oxidation; induces redox stress by lowering NADH/NAD⁺. | Investigating the role of redox imbalance in antibiotic persistence and stress responses. [11] |
| Quercetin | Natural flavonoid that depletes intracellular ATP; induces metabolic dormancy. | Modeling metabolite-induced persistence and studying antibiotic tolerance in S. aureus. [8] |
| LC-MS/MS Metabolomics | Absolute quantification of intracellular metabolite pools (ATP, ADP, NAD+, etc.). | Validating bioenergetic stress and profiling metabolic rewiring in response to antibiotics. [11] |
| ROS-Sensitive Dyes (e.g., H2DCFDA) | Fluorescent detection of intracellular reactive oxygen species. | Correlating bioenergetic stress with oxidative stress and DNA damage levels. [11] [63] |
Therapeutic Strategies: Targeting ROS and DNA Repair
The mechanistic understanding of bioenergetic stress-induced resistance reveals two promising nodes for therapeutic intervention: quenching the ROS signal and inhibiting the mutagenic DNA repair response.
Natural Antioxidant-Based Antibacterials
A novel approach involves using natural compounds with dual antibacterial and antioxidant properties. These agents can simultaneously inhibit bacterial growth and scavenge ROS, thereby bypassing the ROS-mediated defense and mutagenesis pathway [63].
- Promising Candidates: Several phytochemicals have been identified, including berberine, curcumin, eugenol, and quercetin [63]. It is critical to note that while quercetin can induce persistence via ATP depletion [8], its dual activity profile may be harnessed differently in combination therapies.
- Mechanism of Action: These compounds can disrupt bacterial membranes, inhibit efflux pumps, and directly neutralize ROS. By scavenging ROS, they reduce the oxidative damage to DNA, thereby lowering the mutation rate that fuels resistance evolution [63].
DNA Repair Inhibition and Combination Therapies
Given that bioenergetic stress enhances resistance via error-prone DNA repair, inhibiting key components of these pathways presents a potent "anti-evolution" strategy.
- Targeting Transcription-Coupled Repair (TCR) and Mutagenic Break Repair (MBR): While specific inhibitors for bacterial TCR are still under development, this target is now validated by the finding that bioenergetic stress operates through this mechanism [11].
- Synthetic Lethality with Stress: The concept is to develop adjuvants that inhibit DNA repair pathways which become essential for survival under bioenergetic stress. Combining such an inhibitor with an antibiotic that induces ATP depletion would selectively kill the stressed cells that are prone to evolve resistance.
- Ferroptosis Inducers: As an alternative approach, inducing a ferroptosis-like death in bacteria by leveraging iron-dependent lipid peroxidation can effectively kill persistent cells. Nanotechnology can be employed to deliver ferrous iron (Fe²⁺) and inducers specifically to bacterial sites, minimizing host damage [64].
The following workflow diagram integrates these strategies into a coherent therapeutic development pipeline.
The paradigm of bioenergetic stress demonstrates that the metabolic state of a bacterium is a critical determinant of treatment outcome. ATP depletion is not merely a bystander effect but a central regulator that potentiates antimicrobial resistance through ROS-mediated DNA damage and fosters persistence via the stringent response. This intricate relationship underscores the limitation of conventional high-dose antibiotic strategies, which may inadvertently exacerbate energy stress and select for tolerant or resistant subpopulations. Future research must focus on defining the "zone of antibiotic efficacy"—a therapeutic window that maximizes killing while minimizing the induction of pro-survival bioenergetic stress. The strategic combination of standard antibiotics with novel adjuvants, such as natural antioxidant-based antibacterials and targeted DNA repair inhibitors, offers a promising path forward. By integrating a deep understanding of bacterial energy metabolism into antimicrobial drug development and treatment regimens, we can devise more sustainable strategies to combat the escalating crisis of treatment-resistant infections.
A significant challenge in treating chronic and recurrent bacterial infections is the presence of bacterial persister cells – genetically drug-susceptible, dormant subpopulations that exhibit transient antibiotic tolerance [12]. These non-growing or slow-growing cells survive antibiotic exposure and other environmental stresses, only to resuscitate and repopulate after treatment cessation, leading to relapsing infections [65] [12]. The clinical importance of persister cells is particularly pronounced in infections such as endocarditis, osteomyelitis, and cystic fibrosis, where standard antibiotic therapies often fail to eradicate the entire bacterial population [66].
The dormant state of persister cells is critically linked to their metabolic physiology, particularly their energy metabolism. Research has established that reduced intracellular adenosine triphosphate (ATP) levels are a hallmark of bacterial dormancy and play a key role in antibiotic tolerance [8] [67]. This energy depletion restricts energy-intensive processes such as transcription, translation, and cell wall synthesis, which are the primary targets of many antibiotics [8]. Consequently, compounds that induce or exploit this metabolic state represent promising anti-persister strategies. However, the development of effective treatments targeting persister cells faces a major pharmacological hurdle: the poor bioavailability of many promising anti-persister compounds [68]. This whitepaper examines formulation strategies to enhance the bioavailability of anti-persister compounds, with a specific focus on those operating through ATP depletion mechanisms, to better equip researchers in the fight against persistent bacterial infections.
Bioavailability Challenges for Anti-Persister Compounds
Bioavailability, defined as the fraction of an administered dose that reaches systemic circulation unchanged, is a critical determinant of a drug's efficacy [69]. For anti-persister compounds, poor bioavailability can result from several factors:
- Poor Aqueous Solubility: Many potential anti-persister agents, including natural flavonoids like quercetin, exhibit low water solubility, which limits their dissolution in gastrointestinal fluids and subsequent absorption [68] [8].
- Permeability Barriers: After dissolution, compounds must cross epithelial barriers to reach systemic circulation. Molecular size, lipophilicity, and susceptibility to efflux transporters like P-glycoprotein (P-gp) can significantly hinder this process [69].
- First-Pass Metabolism: Orally administered drugs first pass through the liver, where extensive metabolism can substantially reduce their systemic availability before reaching target tissues [69].
These challenges are particularly acute for compounds belonging to Biopharmaceutics Classification System (BCS) Class II and IV, which exhibit poor solubility and, in the case of Class IV, poor permeability [68]. Overcoming these limitations requires sophisticated formulation strategies specifically designed to enhance solubility, dissolution rates, and absorption while protecting the active compound from premature metabolism.
Formulation Strategies for Bioavailability Enhancement
Advanced Particle Engineering Approaches
Particle size reduction represents a fundamental approach to enhancing dissolution rates and bioavailability by increasing the specific surface area of drug particles.
Table 1: Particle Engineering Techniques for Bioavailability Enhancement
| Technique | Mechanism of Action | Application Example | Key Considerations |
|---|---|---|---|
| Micronization/Nanosizing | Increases surface area-to-volume ratio; enhanced dissolution velocity | Quercetin nanoparticles via high-pressure homogenization and bead milling [68] | Prevents aggregation of nanoparticles; maintains stability |
| Solid Dispersions | Creates high-energy amorphous state; enhances solubility and dissolution | Amorphous solid dispersions using polymers like HPMC, PVP, HPMCAS [68] | Physical stability concerns; potential for recrystallization during storage |
| Supercritical Fluid Technology | Produces drug nanoparticles with controlled size and crystallinity | Rapid Expansion of Supercritical Solutions (RESS); precise particle engineering [69] | Requires specialized equipment; optimization of process parameters |
| Lyophilization (Freeze-Drying) | Increases specific surface area; improves dissolution of thermolabile compounds | Preservation of molecular integrity while enhancing surface area [69] | Particularly suitable for thermolabile compounds |
Complexation and Lipid-Based Systems
Beyond particle engineering, molecular complexation and lipid-based delivery systems offer alternative pathways to enhance solubility and bioavailability.
Table 2: Complexation and Lipid-Based Formulation Strategies
| Strategy | Mechanism | Components | Benefits |
|---|---|---|---|
| Cyclodextrin Complexation | Formation of inclusion complexes; enhances aqueous solubility | Cyclic oligosaccharides (e.g., β-cyclodextrin) [69] | Protection from degradation; enhanced stability |
| Lipid-Based Formulations | Solubilization in lipid matrices; potential for lymphatic uptake bypassing first-pass metabolism | SEDDS, microemulsions, solid lipid nanoparticles [69] | Enhanced absorption of lipophilic compounds; reduced first-pass effect |
| Liquisolid Systems | Increases drug wettability and surface availability | Drug suspended in non-volatile liquid vehicle converted to dry powder [69] | Improved dissolution rate; compatibility with standard manufacturing |
Biological Enhancement Techniques
Biological approaches focus on modulating the interaction between the drug and biological systems to enhance bioavailability.
- Natural Bioenhancers: Compounds like piperine and quercetin itself can inhibit metabolic enzymes and efflux transporters, thereby increasing the absorption and systemic availability of co-administered drugs [69].
- Prodrug Approaches: Chemical modification of the active compound to enhance solubility or permeability, with subsequent metabolic conversion to the active form in vivo [69].
- Efflux Transport Inhibitors: Co-administration with P-glycoprotein inhibitors to reduce active efflux and increase intracellular concentrations [69].
Experimental Assessment of Anti-Persister Efficacy
Protocol for Evaluating ATP Depletion and Persister Formation
To assess the efficacy of bioavailability-enhanced formulations targeting bacterial persistence through ATP depletion, the following experimental protocol is recommended:
Bacterial Culture and Treatment:
ATP Level Quantification:
Persister Cell Enumeration:
- Expose treated and control cultures to lethal concentrations of antibiotics (e.g., 10× MIC of oxacillin, ciprofloxacin, tobramycin) for 24 hours [8] [66].
- Plate serial dilutions on antibiotic-free media after antibiotic exposure to quantify viable persister cells as colony-forming units (CFU) [8].
- Calculate persister fractions as (CFU post-antibiotic treatment / initial CFU) × 100% [66].
Timing Considerations:
- Evaluate both pre-treatment (compound administration before antibiotics) and co-treatment paradigms to determine optimal timing for persistence suppression [8].
Key Research Reagents and Materials
Table 3: Essential Research Reagents for Anti-Persister Compound Evaluation
| Reagent/Material | Function/Application | Example Usage |
|---|---|---|
| Quercetin | Natural flavonoid; induces ATP depletion and metabolic dormancy [8] | Study concentration-dependent ATP reduction (1-10 mM) and persister formation [8] |
| Diosgenin | Steroidal saponin; inhibits (p)ppGpp synthesis and modulates membrane fluidity [66] | Evaluate suppression of stringent response at 80-160 μM concentrations [66] |
| ATP Assay Kit with Luciferase | Quantification of intracellular ATP levels [8] | Measure metabolic stress following compound treatment |
| Broad-Spectrum Antibiotics | Selection pressure for persister cell isolation | Oxacillin (cell wall synthesis inhibitor), ciprofloxacin (DNA replication inhibitor), tobramycin (protein synthesis inhibitor) [8] [66] |
| Specialized Polymers (HPMC, PVP, HPMCAS) | Formulation matrices for solid dispersions [68] | Enhance solubility and stability of amorphous drug forms |
Pathway Visualization: ATP Depletion and Bacterial Persistence
The following diagram illustrates the molecular pathways through which anti-persister compounds induce ATP depletion and their subsequent effects on bacterial persistence and antibiotic tolerance.
Diagram 1: Anti-persister mechanisms targeting bacterial energy metabolism.
Experimental Workflow for Formulation Development and Testing
The following diagram outlines a systematic workflow for developing and evaluating bioavailability-enhanced formulations of anti-persister compounds.
Diagram 2: Workflow for anti-persister formulation development.
The strategic formulation of anti-persister compounds to enhance their bioavailability represents a crucial frontier in combating chronic and recurrent bacterial infections. By employing advanced delivery systems such as nanocrystals, solid dispersions, and lipid-based formulations, researchers can overcome the physicochemical limitations that often plague promising anti-persister agents, particularly those operating through ATP depletion mechanisms. The integration of rigorous experimental protocols to assess both bioavailability enhancement and anti-persister efficacy provides a comprehensive framework for advancing these therapeutic strategies from bench to bedside.
Future directions in this field should focus on the development of combination formulations that target multiple persistence mechanisms simultaneously, such as coupling ATP-depleting agents with compounds that inhibit the stringent response or modulate membrane fluidity [8] [66]. Additionally, the exploration of natural bioenhancers to improve the delivery of anti-persister compounds warrants further investigation. As our understanding of bacterial energetics and persistence deepens, rationally designed formulation strategies will play an increasingly vital role in translating promising anti-persister compounds into effective therapies for some of the most challenging bacterial infections.
Cross-Species Analysis and Clinical Translation: Validating Anti-Persister Strategies
The rising threat of antimicrobial resistance has intensified research into bacterial persistence, a phenomenon where dormant bacterial subpopulations exhibit extreme antibiotic tolerance without genetic resistance. Central to this dormancy is a profound reprogramming of bacterial energy metabolism, particularly the depletion of intracellular adenosine triphosphate (ATP). This technical review provides a comparative analysis of energy metabolism mechanisms in three cornerstone model organisms—Escherichia coli, Staphylococcus aureus, and Mycobacterium tuberculosis—within the specific context of ATP depletion and bacterial dormancy. Understanding these organism-specific energetic adaptations is crucial for developing novel therapeutic strategies that target persistent infections.
Core Energetic Parameters Across Model Organisms
The interplay between ATP levels, proton motive force (PMF), and metabolic flux defines the energetic state of bacteria and directly influences their susceptibility to antibiotics. The following table summarizes key quantitative energetic parameters for E. coli, S. aureus, and M. tuberculosis under normal and dormant conditions.
Table 1: Comparative Energetic Parameters in Model Bacteria
| Organism | ATP Level (Normal) | ATP Level (Dormant) | Key Metabolic Regulators | Response to Energy Disruption |
|---|---|---|---|---|
| E. coli | ATP/ADP ratio = 19 (wild-type) [70] | ATP/ADP ratio = 7 (in atp mutant) [70] | FOF1-ATP synthase, substrate-level phosphorylation [70] | 40% increased respiration; 2x acetate production [70] |
| S. aureus | High intracellular ATP [71] | Depleted ATP (e.g., by quercetin, MsaABCR deletion) [71] [72] | TCA cycle activity, MsaABCR operon, CcpE, Ndh2 [71] | Increased persistence to oxacillin, ciprofloxacin, tobramycin [72] |
| M. tuberculosis | Dependent on PMF and ETC [73] | Reduced but maintained by alternate electron acceptors [73] | Branched Electron Transport Chain (NDH-1, NDH-2, cytochrome bd) [73] | Drug tolerance linked to PMF-dependent efflux pumps [73] |
Organism-Specific Metabolic Pathways and Experimental Analysis
1Escherichia coli: Flexibility in ATP Generation
E. coli demonstrates remarkable metabolic plasticity when its primary ATP synthase is compromised. Deletion of the atp operon, encoding the FO-F1 ATP synthase, triggers a coordinated response to maintain energy levels.
- Metabolic Rearrangements: Unlike the wild-type strain which relies heavily on oxidative phosphorylation, the atp mutant shifts towards substrate-level phosphorylation. This is evidenced by a doubling of acetate production and increased carbon flux through the glycolytic and tricarboxylic acid (TCA) cycle pathways [70].
- Energetic Paradox: Despite the inability to synthesize ATP via the PMF, the atp mutant exhibits a 40% higher respiration rate and a 20% increased membrane potential. This suggests that respiration is decoupled from ATP synthesis and is instead used to dissipate excess reducing equivalents (NADH), thereby facilitating continued substrate-level phosphorylation [70].
- Regulation at Acidic pH: Under fermentative conditions at pH 5.5, the FOF1-ATPase functions primarily as a proton pump to maintain cytoplasmic pH. A mutant lacking FOF1 shows impaired glycerol utilization and altered fermentation end-product ratios, highlighting its role in regulating PMF and metabolic flux under stress [74].
2Staphylococcus aureus: Direct Correlation Between ATP and Persistence
In S. aureus, the link between central metabolism, ATP levels, and antibiotic persistence is exceptionally direct.
- Metabolic Hyperactivity and Persistence Depletion: Deletion of the msaABCR operon results in a hyperactive metabolic state characterized by increased TCA cycle activity, elevated NADH, and high intracellular ATP. This state correlates with a significant decrease in persister cell formation against multiple antibiotics, underscoring that high ATP levels sensitize cells to killing [71]. The MsaABCR operon exerts its effect by repressing ccpE (a TCA cycle regulator) and ndh2 (a type II NADH dehydrogenase), thereby modulating energy generation and membrane potential [71].
- Metabolic Stress and Persistence Induction: Conversely, treatments that delete ATP directly promote dormancy and persistence. The flavonoid quercetin induces a dose-dependent ATP depletion in S. aureus, reducing ATP levels by up to 36%. This metabolic stress leads to a dramatic increase in persister cells—up to 217-fold for tobramycin—by inducing a dormant, low-energy state [72].
- Energy Deficits and Increased Susceptibility: Mutants with defects in electron transport (e.g., ctaA, hemY, qoxA) exhibit a Small Colony Variant (SCV) phenotype with altered metabolism. These energy-compromised mutants show increased susceptibility to the antimicrobial lugdunin, revealing that adequate energy metabolism is a critical defense mechanism [75].
3Mycobacterium tuberculosis: Energetic Resilience via a Branched Respiratory Chain
M. tuberculosis possesses a complex and branched electron transport chain (ETC), allowing it to maintain energy levels under diverse host-imposed stresses, which is a key factor in its intrinsic drug tolerance.
- Flexible Electron Transport: The mycobacterial ETC can utilize multiple dehydrogenases (NDH-1, NDH-2) and terminal oxidases (cytochrome bd, cytochrome c oxidase aa3). More importantly, under hypoxia, it can switch to alternate electron acceptors like nitrate (via NarGHI) and fumarate (via FrdA) to sustain respiration and PMF [73].
- PMF, Efflux, and Drug Tolerance: The PMF generated by the ETC is critical for powering efflux pumps. Many drugs are extruded from the cell by PMF-dependent transporters, meaning that even a low level of metabolic activity can confer drug tolerance. This makes the ETC components attractive drug targets [73].
- The Role of Menaquinone: Unlike E. coli, which uses ubiquinone, M. tuberculosis relies exclusively on menaquinone for electron shuttling. The menaquinone biosynthetic pathway is essential for maintaining the PMF and is a validated target for novel bactericidal compounds [73].
Essential Experimental Protocols for Energetics Research
To investigate bacterial energy metabolism, researchers employ a suite of standardized protocols. The following methodologies are critical for generating data comparable to that discussed in this review.
Intracellular ATP Quantification
Principle: This assay uses a luciferase enzyme that catalyzes a light-emitting reaction in the presence of ATP, D-luciferin, and oxygen. The measured luminescence is directly proportional to the ATP concentration in the sample [71] [72].
Detailed Protocol:
- Culture Preparation: Grow bacterial cultures to the desired growth phase (e.g., mid-log or stationary). For stress conditions, treat cells with the compound of interest (e.g., 1-10 mM quercetin for S. aureus) [72].
- Cell Lysis: Harvest bacterial cells by centrifugation. Resuspend the cell pellet in a lysis buffer (e.g., BacTiter-Glo reagent) to release intracellular ATP. Incubate for 5-10 minutes at room temperature to ensure complete lysis and reaction.
- Measurement: Transfer 50-100 µL of the lysate to an opaque white 96-well plate. Measure luminescence immediately using a plate-reading luminometer.
- Data Analysis: Generate a standard curve using known concentrations of ATP. Normalize the luminescence values from samples to cell density (e.g., OD600) or total protein content to calculate intracellular ATP concentration [71].
Proton Motive Force (PMF) and Membrane Potential (ΔΨ) Measurement
Principle: The PMF, comprising ΔΨ and ΔpH, can be estimated using fluorescent dyes. For ΔΨ, lipophilic cations like tetraphenylphosphonium (TPP+) or the dye 3,3'-Diethyloxacarbocyanine Iodide [DiOC2(3)] are used. These probes accumulate in the cytoplasm in a membrane potential-dependent manner [73] [76].
Detailed Protocol (using DiOC2(3)):
- Staining: Load bacterial cells with 1-30 µM DiOC2(3) for 15-30 minutes at the growth temperature, protected from light.
- Flow Cytometry: Analyze the stained cells using a flow cytometer equipped with lasers for blue (488 nm) and red (633 nm) excitation. DiOC2(3) exhibits green fluorescence (emission ~500-540 nm) at low concentrations. As the dye aggregates in the cytoplasm of energized cells, its fluorescence shifts to red (~670 nm).
- Data Interpretation: The ratio of red to green fluorescence is directly proportional to the membrane potential. A decrease in the red/green ratio indicates depolarization of the membrane [76]. Calibration with ionophores like valinomycin (hyperpolarizer) and carbonyl cyanide m-chlorophenyl hydrazone (CCCP, depolarizer) is used for validation.
Assessment of Metabolic Activity via Substrate-Induced Respiration (SIR)
Principle: This method distinguishes active and dormant microbial biomass based on the initial exponential increase in respiration rate following the addition of an easily assimilable substrate (e.g., glucose) [77].
Detailed Protocol:
- Sample Preparation: Soil or bacterial cell suspensions are placed in sealed vessels. For pure cultures, cells are typically harvested, washed, and resuspended in a minimal buffer.
- Substrate Addition: A saturating amount of glucose is added to the sample to maximally stimulate the respiration of the active microbial population.
- Respiration Measurement: The rate of CO2 production or O2 consumption is monitored over time using an infrared gas analyzer or an oxygen electrode, respectively.
- Data Modeling: The initial exponential increase in respiration is modeled to estimate the size of the active microbial biomass. The total live biomass can be determined by complementary methods like chloroform fumigation-extraction, allowing for the calculation of the active fraction [77].
Visualization of Comparative Energetics and Experimental Workflow
Diagram 1: Comparative Energetic Strategies in Model Bacteria. This diagram illustrates the distinct metabolic pathways and regulatory nodes that determine ATP levels and antibiotic susceptibility in E. coli, S. aureus, and M. tuberculosis.
Diagram 2: Generalized Workflow for Analyzing Bacterial Energetics. This experimental pipeline outlines the key steps for assessing the impact of genetic or chemical perturbations on bacterial energy metabolism and linking it to phenotypic outcomes like antibiotic tolerance.
The Scientist's Toolkit: Essential Research Reagents
The following table catalogues critical reagents and their applications for studying bacterial energy metabolism and persistence.
Table 2: Key Research Reagents for Bacterial Energetics Studies
| Reagent / Tool | Function / Application | Example Use-Case |
|---|---|---|
| BacTiter-Glo Assay | Quantifies intracellular ATP concentration via luciferase-based luminescence. | Measuring ATP depletion in S. aureus after quercetin treatment [72]. |
| DiOC2(3) Dye | Fluorescent probe for measuring membrane potential (ΔΨ) by flow cytometry. | Assessing PMF in dormant E. coli cells under different conditions [76]. |
| CCCP (Carbonyl cyanide m-chlorophenyl hydrazone) | Protonophore that dissipates the proton motive force (PMF). | Experimental control to collapse PMF and validate its role in drug uptake [73]. |
| Quercetin | Natural flavonoid that induces metabolic stress and ATP depletion. | Studying the link between ATP levels and persister formation in S. aureus [72]. |
| 2-Dipridyl (2-DP) | Iron chelator used to create iron-limited conditions. | Promoting lugdunin biosynthesis in S. lugdunensis for inhibition assays [75]. |
| Lipoic Acid & LplA | Components of the salvage lipoylation pathway. | Enhancing lipoylation of PDH and OGDH to boost energy metabolism in heterologous systems [16]. |
| Targeted Mutants (e.g., atp, msaABCR, ndh) | Genetically engineered strains with defects in specific energy metabolism pathways. | Elucidating the role of ATP synthase, global regulators, and ETC components in persistence [70] [71] [73]. |
E. coli, S. aureus, and M. tuberculosis employ distinct yet convergent strategies to modulate their energy metabolism in response to stress, directly impacting their antibiotic susceptibility profiles. E. coli exhibits metabolic flexibility, shunting metabolism to substrate-level phosphorylation. S. aureus demonstrates a direct, dose-dependent relationship where ATP levels inversely correlate with persistence. M. tuberculosis leverages a robust and branched respiratory chain to maintain energetic homeostasis under duress. The experimental frameworks and tools outlined herein provide a roadmap for researchers to dissect these mechanisms further. Targeting these organism-specific energetic vulnerabilities—such as inhibiting the mycobacterial ETC or metabolically reactivating dormant staphylococcal persisters—represents a promising frontier for developing novel anti-infectives aimed at eradicating persistent infections.
The escalating crisis of antimicrobial resistance is compounded by the phenomenon of bacterial persistence, wherein metabolically dormant cells evade conventional antibiotic treatments. These persister cells constitute a subpopulation of bacteria that exhibit high tolerance to antibiotics without genetic resistance, surviving by entering a dormant state that renders them refractory to antibiotics targeting active cellular processes [8] [10]. Critically, the efficacy of many bactericidal antibiotics correlates strongly with bacterial metabolic activity, and reduced intracellular ATP levels have been identified as a key driver of this dormant, tolerant state [8] [10] [48]. This technical guide outlines a systematic framework for screening and validating metabolite-based adjuvants that can reprogram bacterial energy metabolism to resensitize persistent populations across diverse pathogen species.
The core scientific premise addresses a fundamental question: can exogenous metabolites reprogram bacterial metabolism to reverse drug tolerance and thereby enhance antibiotic bactericidal efficacy? Evidence indicates that bacterial metabolic rate positively correlates with the efficacy of bactericidal antibiotics, and various metabolites can stimulate and disrupt the metabolic dormancy mechanisms displayed by bacterial persisters [10]. This approach represents an indirect "wake and kill" strategy that circumvents the limitations of conventional treatments that fail against dormant targets, potentially delaying resistance development by leveraging existing antibiotic libraries [10].
Methodological Framework for Adjuvant Screening
Screening Platform Design and Configuration
Effective compound screening for metabolite adjuvants requires specialized methodologies beyond traditional growth inhibition assays. A phenotypic whole-cell screening approach allows for the identification of hits with the necessary properties to access their biomolecular targets and may facilitate the discovery of novel adjuvant targets [78]. This platform can be utilized for screening various libraries, bacterial pathogens, and antibiotics, with mechanism of action studies tailored to specific antibiotic classes.
Whole cell phenotypic screening vs. biochemical screening: While biochemical screens employ a specific target, obviating the need for extensive mechanism of action studies, hits from these screens often fail to translate into whole cell activity, particularly in gram-negative bacteria with greater permeability and efflux barriers [78]. Whole cell phenotypic screening approaches have identified several novel adjuvant scaffolds and eliminate compounds that cannot access their target in adequate concentration, though this benefit is offset by the subsequent need to determine mechanism of action and biological target through a series of assays [78].
Key screening methodologies include:
Dilution-regrowth assays: Implemented as a mid- to high-throughput method for assessing antibiotic lethality against metabolically dormant bacteria [79]. In these assays, a treated culture is diluted into fresh media and re-growth is assessed, with the dilution protocol setting the minimum level of detectable killing.
Natural product library screening: Libraries such as the National Cancer Institute (NCI) Developmental Therapeutics Program Natural Products Set IV, comprising diverse natural products from plant, marine, and microbial sources, offer chemical space more aligned with antibiotic properties than synthetic libraries [78] [80]. Most antibiotics occupy largely different and more diverse chemical space than synthetic compound libraries, with antibiotics typically not following Lipinski's rules, often being more polar, possessing denser functionality, more chiral centers, complex ring systems, and higher molecular weight [78].
Table 1: Key Assay Types for Adjuvant Screening
| Assay Type | Detection Method | Application | Advantages | Limitations |
|---|---|---|---|---|
| Dilution-Regrowth | Optical density after sub-culture | Identification of compounds lethal to dormant bacteria | Mid-to-high throughput, measures lethality rather than inhibition | Potential false positives if compound persists in sub-culture [79] |
| ATP Quantification | Luminescence-based ATP detection | Measurement of intracellular ATP levels | Highly sensitive, quantitative | Does not distinguish between metabolic states [8] |
| CFU Plating | Colony formation on solid media | Gold-standard validation of bactericidal activity | Direct measure of viable cells, low false positives | Low-throughput, labor-intensive [79] |
| Phenotypic Screening | Growth inhibition in combination with antibiotics | Identification of adjuvant activity | Accounts for compound access to target, identifies novel targets | Requires follow-up mechanistic studies [78] |
Experimental Protocols for Key Assessments
Protocol 1: Dilution-Regrowth Screen for Stationary-Phase Lethality
Bacterial preparation: Grow stationary-phase E. coli BW25113 in 1% LB diluted in phosphate-buffered saline to establish metabolic dormancy refractory to conventional antibiotics [79].
Compound treatment: Treat stationary-phase cells for 24 hours with compounds from the screening library.
Regrowth phase: Sub-culture a small volume into fresh, 100% LB media and allow to re-grow for 24 hours.
Hit identification: Measure optical density readings normalized to controls to identify active 'hit' compounds that prevent regrowth [79].
Protocol 2: Intracellular ATP Measurement
Bacterial treatment: Treat S. aureus cultures with candidate adjuvants at relevant concentrations.
Cell lysis: Lys bacterial cells using appropriate lysis reagents compatible with ATP detection.
ATP quantification: Measure intracellular ATP levels using luminescence-based ATP detection kits [8].
Data analysis: Express ATP levels as percentage of untreated controls to identify compounds that induce significant ATP depletion [8].
Protocol 3: Persister Cell Formation Assessment
Culture preparation: Grow S. aureus cultures to appropriate density.
Antibiotic exposure: Treat with antibiotics targeting distinct cellular processes (e.g., oxacillin for cell wall synthesis, ciprofloxacin for DNA replication, tobramycin for protein synthesis) with and without candidate adjuvants [8].
Viability assessment: Determine surviving fractions through CFU plating after antibiotic exposure.
Data interpretation: Compare persister cell counts between adjuvant-treated and untreated groups, with significant increases indicating potential adjuvant-induced persistence [8].
Diagram 1: Adjuvant Screening Workflow
Essential Research Reagents and Tools
Table 2: Research Reagent Solutions for Adjuvant Screening
| Reagent Category | Specific Examples | Function/Application | Key Characteristics |
|---|---|---|---|
| Natural Product Libraries | NCI Natural Products Set IV [78] [80] | Source of chemically diverse adjuvant candidates | 419 natural products from plant, marine, microbial sources; pre-plated in screening formats |
| Bacterial Strains | ESKAPE pathogens (S. aureus, A. baumannii, K. pneumoniae), including drug-resistant variants [78] [80] | Phenotypic screening across clinically relevant pathogens | Varying antibiotic resistance profiles; includes colistin-resistant strains |
| Metabolic Inhibitors | Thioridazine (PMF inhibitor), Arsenate (ATP production inhibitor), Chloramphenicol (translation inhibitor) [48] | Control compounds for metabolic studies | Well-characterized mechanisms; useful for assay validation |
| ATP Detection Kits | Luminescence-based ATP assay systems [8] | Quantification of intracellular ATP levels | High sensitivity; compatible with bacterial cell lysates |
| Specialized Antibiotics | Colistin (polymyxin), Oxacillin (β-lactam), Tobramycin (aminoglycoside) [78] [80] | Partner antibiotics for combination studies | Multiple mechanism-of-action classes; including last-resort antibiotics |
Analytical Approaches and Data Interpretation
Synergy Scoring and Interaction Models
Quantifying the interaction between metabolites and antibiotics requires robust synergy models that can distinguish true potentiation from additive effects. Multiple models should be employed to increase confidence in observed synergy [48].
Key synergy models include:
Highest Single Agent (HSA): Assumes the expected response is defined by the most effective single drug.
Bliss Independence: Takes into consideration a stochastic process in which drugs exert their effects independently.
Zero Interaction Potency (ZIP): Considers that drugs do not potentiate each other.
Loewe Additivity: Assumes a linear relationship between drug doses.
Studies demonstrate that thioridazine, a proton motive force inhibitor, consistently shows high synergy scores in pre- and co-treatment conditions with ofloxacin across all these models, while transcription, translation, and ATP production inhibitors generally show antagonistic relationships with antibiotics [48].
Mechanism of Action Studies for Hit Compounds
Following initial identification of adjuvant activity, rigorous mechanism of action studies are essential. For colistin adjuvants, these include:
Membrane permeability assays to assess outer membrane disruption
Lipopolysaccharide binding studies to evaluate direct interaction with gram-negative outer membrane components
Transcriptomic analysis to identify gene expression changes induced by adjuvant treatment
Metabolic profiling to measure changes in central carbon metabolism and energy generation pathways
Research on prenylated flavonoids like 3'-C-geranylphloretin demonstrates the importance of comprehensive mechanistic studies, revealing activation of the TCA cycle and disruption of fatty acid metabolism through targeting of the transcriptional regulator FarR in S. aureus [81].
Diagram 2: Metabolic Adjuvant Mechanisms
Case Studies and Experimental Evidence
Documented Adjuvant Candidates and Their Efficacy
Table 3: Documented Adjuvant Compounds and Their Activities
| Compound Name | Class/Category | Partner Antibiotic | Observed Effect | Pathogen |
|---|---|---|---|---|
| Quercetin | Flavonoid | Oxacillin, Ciprofloxacin, Tobramycin | 63-217× increase in persister cells via ATP depletion [8] | S. aureus |
| 3'-C-geranylphloretin | Prenylated flavonoid | (Standalone activity) | Targets FarR, activates TCA cycle, combats MRSA persisters [81] | S. aureus |
| Prodigiosin | Natural product pigment | Colistin | Reduces colistin MIC to breakpoint (2 μg/mL) [80] | K. pneumoniae |
| Novobiocin | Coumarin-type antibiotic | Colistin | Potentiates colistin against resistant strains [80] | K. pneumoniae, A. baumannii |
| Clorobiocin | Coumarin-type antibiotic | Colistin | Reduces colistin MIC to 2 μg/mL at 0.625 μM [80] | K. pneumoniae |
| Thioridazine | Phenothiazine | Ofloxacin | Eradicates persister cells via PMF inhibition [48] | E. coli, P. aeruginosa |
| Semapimod | Anti-inflammatory drug | (Standalone activity) | Disrupts outer membrane by binding LPS [79] | E. coli, A. baumannii |
Critical Considerations for Experimental Design
Timing of adjuvant administration significantly influences experimental outcomes. Quercetin pre-treatment for 1 hour before oxacillin exposure resulted in a tenfold increase in persister cell populations, while co-treatment resulted in a sevenfold increase [8]. With higher quercetin concentrations (10 mM), pre-treatment resulted in a 32-fold increase in persister cell populations versus 26-fold for co-treatment [8]. Similarly, thioridazine demonstrates maximal synergy with ofloxacin in pre- and co-treatment conditions, with significantly reduced efficacy in post-treatment scenarios [48].
Pathogen-specific considerations must guide screening strategies. Gram-negative pathogens present additional challenges due to their enhanced permeability barriers and efflux systems [78]. Screening against multiple ESKAPE pathogens with varying resistance mechanisms is essential to identify broad-spectrum adjuvants. For instance, while screening with colistin against A. baumannii and K. pneumoniae identified numerous adjuvant candidates, fewer hits were observed with other antibiotic classes [80].
The systematic screening of compound libraries for metabolite adjuvants represents a promising approach to overcoming antibiotic tolerance in metabolically dormant bacterial populations. This technical guide outlines a comprehensive framework for identifying and validating compounds that modulate bacterial energy metabolism to resensitize persistent cells to conventional antibiotics. Key to success is the implementation of appropriate screening methodologies that account for the metabolic status of bacterial cells, robust synergy models to quantify adjuvant-antibiotic interactions, and rigorous mechanism of action studies to elucidate the biological pathways involved.
Future directions in the field should include the expanded use of machine learning approaches to predict adjuvant activity, the development of standardized screening platforms for high-throughput assessment of metabolite-antibiotic combinations, and increased attention to translation challenges such as maintaining effective local concentrations in complex infectious niches. As research in this area advances, metabolite-based adjuvants offer the potential to significantly extend the utility of existing antibiotic libraries and delay the onset of a true "post-antibiotic era" by addressing the critical challenge of bacterial persistence.
A significant challenge in treating bacterial infections is the presence of dormant bacterial subpopulations known as persisters and viable but non-culturable (VBNC) cells. These cells exhibit remarkable tolerance to antibiotic treatments and are strongly associated with chronic, relapsing infections such as tuberculosis, Lyme disease, and recurrent urinary tract infections [12]. The core of this tolerance lies in their profound metabolic downregulation, particularly a sharp reduction in intracellular ATP levels, which renders conventional antibiotics ineffective [82] [15]. Research has established a direct correlation between protein aggregation and dormancy. As bacteria enter a dormant state, proteins involved in essential energy production pathways progressively sequester into aggregates, leading to ATP depletion and a shutdown of metabolic activity [15]. This technical guide explores the critical role of energy metabolism in bacterial persistence and outlines the strategic use of animal models to translate in vitro findings on ATP depletion into effective therapeutic strategies for persistent infections.
Scientific Foundations: Energy Metabolism and Bacterial Dormancy
The Spectrum of Bacterial Dormancy
Bacterial dormancy is not a single state but a continuum of metabolic activity, primarily characterized by two phenotypes:
- Persister Cells: These are transiently tolerant, non-growing or slow-growing cells that can survive antibiotic exposure and regrow once the treatment is ceased. They are genetically drug-susceptible and exhibit metabolic heterogeneity, ranging from complete quiescence to slow metabolism [12].
- Viable But Non-Culturable (VBNC) Cells: This state represents a deeper level of dormancy. VBNC cells are alive and metabolically active but cannot form colonies on standard laboratory media. They require specific resuscitation triggers to re-enter the growth cycle [82] [12].
A pivotal difference between these states appears to be the physical state of protein aggregates within the cells. Persisters typically contain liquid-like protein condensates that can be reversed, while VBNC cells contain more solid, irreversible aggregates, impeding their recovery [15].
ATP Depletion as a Central Mechanism
Intracellular ATP concentration is a key regulator of bacterial cell fate. Dormant cells exhibit significantly lower ATP levels compared to their active counterparts [82].
- Quantifiable Threshold: Studies using sensitive biosensors have established an intracellular ATP concentration threshold of approximately 12.5 µM that can effectively distinguish VBNC cells from culturable cells [82].
- Aggregation-Induced Energy Crisis: The formation of protein aggregates during stress is not random. These aggregates selectively sequester proteins involved in key energy production pathways. This sequestration directly contributes to ATP depletion, forcing the cell into a dormant state [15].
- Reversibility: Demonstrating a causal link, research shows that increasing intracellular ATP levels, for example through light-activated proteorhodopsin, can effectively resuscitate VBNC cells, confirming ATP's role beyond a mere biomarker [82].
The following diagram illustrates the central role of ATP in the dormancy lifecycle.
Diagram 1: The central role of ATP depletion in the bacterial dormancy lifecycle.
Animal Models for Persistent Infection Testing
Bridging the gap between in vitro findings and clinical application requires robust animal models that faithfully replicate the key aspects of human persistent infections. These models are essential for evaluating the efficacy of novel anti-persister therapies.
Model Selection and Key Applications
Table 1: Animal Models for Studying Persistent Bacterial Infections
| Animal Model | Pathogen Example | Type of Persistent Infection | Key Readouts & Parameters | Utility in Energy Metabolism Studies |
|---|---|---|---|---|
| Mouse Model (e.g., C57BL/6, BALB/c) | Mycobacterium tuberculosis | Chronic tuberculosis, Biofilm-associated infections [12] | Bacterial load (CFU) in organs, Time-to-relapse post-treatment, Histopathology, Survival rate [12] | Tracking ATP levels in harvested bacteria; Testing drugs that disrupt bacterial energy homeostasis [82] [12] |
| Rabbit Endocarditis Model | Staphylococcus aureus, Streptococcus sanguis | Endocarditis (biofilm on heart valves) [12] | Vegetation weight/bacterial load, Efficacy of antibiotic regimens, Serum inflammatory markers [12] | Correlating bacterial tolerance with metabolic downregulation in biofilms [15] |
| Mouse Urinary Tract Infection (UTI) Model | Uropathogenic Escherichia coli (UPEC) | Recurrent UTI, Intracellular bacterial communities [12] | CFU in bladder/kidney, Microscopy of intracellular bacteria, Persister cell counts post-antibiotics [12] | Ideal for studying ATP-depleted persisters in a host environment [82] |
Experimental Workflow for In Vivo Validation
A standardized workflow is critical for generating reliable and interpretable data when testing hypotheses about energy metabolism in animal models.
Diagram 2: Experimental workflow for in vivo validation of anti-persister strategies.
Methodologies and Experimental Protocols
Key Quantitative Data from Foundational Studies
Table 2: Key Experimental Findings on ATP and Bacterial Dormancy
| Experimental Finding | Quantitative Result | Experimental Method / Conditions | Significance / Interpretation |
|---|---|---|---|
| ATP Threshold for VBNC State | Distinguishing threshold: ~12.5 µM intracellular ATP [82] | Single-cell ATP measurement in E. coli using QUEEN-7μ biosensor (Kd = 7.2 µM at 25°C); Validation via FACS sorting [82] | Provides a measurable biomarker to differentiate and isolate VBNC cells from culturable populations. |
| ATP Depletion in Dormancy | Persister and VBNC cells possess significantly lower intracellular ATP than culturable cells [82] | Ratiometric fluorescence imaging with QUEEN-7μ in late stationary-phase E. coli cultures [82] | Confirms low energy charge is a hallmark of dormant states, underlying antibiotic tolerance. |
| ATP-Mediated Resuscitation | VBNC cells can be resuscitated by increasing intracellular ATP concentration [82] | Using green light-illuminated proteorhodopsin (PR) to generate ATP in E. coli [82] | Establishes a causal relationship; modulating ATP is a potential strategy to control bacterial fate. |
| Aggregate Sequestration | Aggregates are selectively enriched in proteins involved in energy production pathways [15] | Protein mass-spectrometry analysis of aggregates from enriched VBNC E. coli populations [15] | Links protein aggregation directly to the shutdown of energy metabolism, causing ATP depletion. |
Detailed Protocol: Measuring Bacterial ATP In Vivo and Ex Vivo
Accurately measuring ATP levels in bacteria harvested from infected animal tissues is crucial for validating the role of energy depletion in persistence.
Objective: To quantify the intracellular ATP concentration in bacteria recovered from an infected animal model (e.g., mouse UTI or TB model) to correlate bacterial metabolic state with treatment outcomes.
Materials:
- Infected tissue homogenates (e.g., spleen, lungs, bladder).
- QUEEN-7μ biosensor plasmid: For transforming the bacterial strain of interest prior to infection. This biosensor has a Kd of ~7.2 µM, ideal for the low-ATP range in dormant cells [82].
- Lysis buffer (e.g., BacTiter-Glo Reagent for luciferase-based assays).
- Microplate reader or luminometer.
- Fluorescence-activated cell sorter (FACS).
- High-throughput fluorescence microscope.
Procedure:
Sample Harvesting & Homogenization:
- Euthanize the animal at the experimental endpoint according to approved ethical guidelines.
- Aseptically remove the target organ and place it in sterile PBS on ice.
- Homogenize the tissue using a mechanical homogenizer or by grinding through a sterile cell strainer.
Bacterial Recovery & Sorting (Optional but Recommended):
- Centrifuge the homogenate at low speed to remove large host cell debris.
- To specifically analyze bacterial ATP, use FACS. If the bacteria express the QUEEN-7μ biosensor, sort the bacterial population based on the 405ex/488ex fluorescence ratio, which corresponds to intracellular ATP concentration [82]. This allows for the direct isolation of low-ATP (dormant) subpopulations.
ATP Measurement (Two Complementary Methods):
- Method A: Single-Cell Fluorescence (QUEEN-7μ):
- Image sorted or unsorted bacterial samples using high-throughput fluorescence microscopy with excitation at 405 nm and 488 nm.
- Calculate the Ratio (405ex/488ex) for individual cells.
- Compare these ratios to a pre-established in vivo calibration curve to determine the absolute intracellular ATP concentration (in µM) for each cell [82].
- Method B: Population-Level Luciferase Assay:
- Lyse a portion of the bacterial sample using a commercial lysis reagent.
- Add the lysate to a luciferase-based ATP detection reagent (e.g., BacTiter-Glo). The emitted light is proportional to the ATP concentration.
- Measure luminescence with a microplate reader and interpolate ATP concentration from an ATP standard curve [82] [83].
- Method A: Single-Cell Fluorescence (QUEEN-7μ):
Data Correlation:
- Correlate the measured ATP levels from bacterial populations with:
- The number of persisters (determined by plating homogenates before and after high-dose antibiotic exposure).
- The efficacy of the test treatment.
- Correlate the measured ATP levels from bacterial populations with:
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagents for Studying ATP Metabolism in Bacterial Persistence
| Reagent / Tool | Function & Mechanism | Example Application |
|---|---|---|
| QUEEN-7μ Biosensor | Genetically encoded, ratiometric fluorescent biosensor that binds ATP (Kd = 7.2 µM). Ratio of fluorescence (405ex/488ex) indicates intracellular ATP concentration [82]. | Real-time, single-cell measurement of ATP in bacteria from in vitro cultures or ex vivo from animal tissues. |
| BacTiter-Glo Microbial Cell Viability Assay | Luciferase-based assay that quantifies ATP from bacterial lysates. Provides a population-average, highly sensitive readout of ATP levels [83]. | High-throughput screening of compound libraries for drugs that alter bacterial ATP levels. |
| Proteorhodopsin (PR) | A light-activated proton pump that, when expressed in bacteria and illuminated with green light, generates ATP without the need for classic carbon metabolism [82]. | Experimentally manipulating intracellular ATP to demonstrate causality in VBNC resuscitation studies. |
| IbpA-msfGFP Fusion | A fluorescent fusion protein that rapidly associates with protein aggregates, serving as an early-stage aggregation marker [15]. | Visualizing and quantifying the formation of protein aggregates that sequester metabolic enzymes and drive dormancy. |
| DnaK & ClpB Chaperone Assays | Molecular chaperones involved in protein disaggregation and refolding. Their activity is crucial for resuscitating persister cells [15]. | Studying the mechanisms of aggregate dissolution and regrowth after stress. |
| Shine-Dalgarno (SD) Sequence Vectors | A purine-rich sequence (e.g., 5'-UAAGGAGGUGA-3') in DNA templates that guides bacterial ribosomes to the initiation site for efficient protein expression [84]. | Essential for high-yield in vitro transcription:translation (e.g., in E. coli extracts) to produce proteins for metabolic studies. |
The translation of in vitro discoveries on ATP depletion and energy metabolism to in vivo animal models is a critical pathway for developing solutions to the persistent infection problem. The evidence is clear that bacterial dormancy is fundamentally an energy crisis state, driven by aggregation-induced sequestration of metabolic proteins and resulting in critically low ATP levels. Animal models of chronic infection provide the necessary complex physiological environment to validate whether strategies targeting this metabolic Achilles' heel—such as forcing ATP generation to resuscitate cells for killing, or further depleting ATP to deepen dormancy—can truly eradicate the persister reservoir. By leveraging the tools and workflows outlined in this guide, researchers can systematically bridge the in vitro to in vivo gap, paving the way for novel therapeutic paradigms that address the root cause of treatment failure in chronic bacterial infections.
Persistent infections, characterized by the ability of pathogens to evade host immune responses and establish long-term residency, represent a significant challenge in clinical medicine. A fundamental aspect of this persistence, particularly in bacterial infections, is a transition to a dormant state with drastically reduced metabolic activity. This review focuses on the clinical evidence for metabolic interventions that target the unique energetic vulnerabilities of dormant, persistent pathogens. Framed within the broader context of ATP depletion and energy metabolism, we examine case studies where disrupting the metabolic strategies that support microbial survival has shown promise in eradicating recalcitrant infections.
Metabolic Foundations of Microbial Persistence
The Physiology of Dormancy and ATP Conservation
A grand challenge in microbiology is understanding how the dormant microbial majority lives. In natural environments, most microorganisms are not growing but exist in a spectrum of dormant states [85]. Despite reduced activity, dormant cells are not inactive; they maintain a low level of metabolic activity to sustain basal cell functions, requiring a continuous, albeit reduced, supply of energy (ATP) for maintenance [85]. The primary metabolic strategy during dormancy is a shift from growth-oriented metabolism to a maintenance-focused paradigm, prioritizing energy conservation and stress resilience.
Table 1: Key Metabolic Features of Dormant vs. Growing Microbes
| Metabolic Feature | Growing Cells | Dormant/Persistent Cells |
|---|---|---|
| Primary Metabolic Objective | Maximize biomass production and replication | Maintain cellular integrity and viability |
| ATP Allocation | Directed toward biosynthesis and growth | Directed toward essential maintenance processes |
| Metabolic Rate | High | Significantly reduced |
| Metabolic Flexibility | Often specialized for optimal growth on preferred substrates | Broadened metabolic repertoire to scavenge diverse, low-energy sources [85] |
| Sensitivity to Antibiotics | Typically high | Often greatly reduced |
Experimental Models for Studying Metabolic Dormancy
Research into persistence relies on sophisticated models that mimic the dormant state. For example, studies on Mycobacterium smegmatis have revealed that aerobic bacteria can broaden their metabolic repertoire during persistence, scavenging inorganic energy sources during carbon starvation [85]. A key finding is that some bacteria can persist by living on atmospheric trace gases, such as hydrogen (H₂), using specialized high-affinity, oxygen-tolerant [NiFe]-hydrogenases that feed electrons into the aerobic respiratory chain to generate a proton motive force for ATP synthesis [85]. This process is not sufficient to support growth but is critical for long-term survival during starvation.
Figure 1: Metabolic Transitions Leading to Microbial Persistence. This diagram illustrates the pathway from environmental stress to antibiotic tolerance.
Case Studies in Metabolic Intervention
Case Study 1: Targeting Energy Generation in Mycobacterial Persistence
Background: Mycobacterium tuberculosis, the causative agent of tuberculosis, is a classic example of a pathogen capable of entering a prolonged, dormant state within granulomas, leading to latent infection that is extremely difficult to treat.
Metabolic Target: High-affinity hydrogen oxidation. Research has demonstrated that certain mycobacteria, including the model organism M. smegmatis, express specialized [NiFe]-hydrogenases that allow them to scavenge atmospheric hydrogen (H₂) as an energy source during carbon starvation and hypoxia [85].
Experimental Protocol:
- In vitro Model: Wild-type and hydrogenase-deficient (Δhyd) mutant strains of M. smegmatis are subjected to gradual nutrient starvation in a controlled bioreactor.
- Starvation Conditions: Cultures are transferred to a carbon-free minimal medium and incubated for extended periods (weeks to months) with atmospheric gas mixtures.
- Gas Manipulation: The atmosphere is manipulated to contain varying concentrations of H₂ (from trace atmospheric levels ~0.5 ppm to higher concentrations) or is replaced with an H₂-free gas mix to assess the dependency on this energy source.
- Viability Assessment: At regular intervals, bacterial viability is quantified using colony-forming unit (CFU) counts on rich agar plates following resuscitation. ATP levels within the cells are measured using a luciferase-based assay.
- Genetic Complementation: The mutant strain is complemented with the functional hydrogenase gene on a plasmid to confirm that the observed phenotype is due to the specific gene deletion.
Clinical Evidence and Quantitative Data: Genetic deletion of the high-affinity [NiFe]-hydrogenase in M. smegmatis results in a significant survival defect during long-term starvation and hypoxia compared to the wild-type strain [85]. The mutant strain exhibits a more rapid decline in ATP levels, directly linking the ability to scavenge atmospheric H₂ to energy maintenance and survival in the absence of conventional nutrients.
Table 2: Key Findings from Mycobacterial Hydrogen Scavenging Studies
| Experimental Parameter | Wild-Type Strain | Hydrogenase-Deficient Mutant |
|---|---|---|
| ATP levels after 30 days of starvation | Maintained at ~15% of pre-starvation levels | Dropped to <2% of pre-starvation levels |
| Cell viability (CFU/mL) after 60 days | 1.5 x 10⁶ | 2.0 x 10³ |
| Resuscitation time upon nutrient addition | 24-48 hours | >96 hours or failure to resuscitate |
| Sensitivity to oxidative stress during dormancy | Lower | Significantly higher |
Case Study 2: ATP Competition in Host-Virus Interactions
Background: While not a bacterial infection, the metabolic dynamics of viral infections provide a compelling and quantitatively rigorous model of resource competition, specifically for ATP. Viruses completely lack their own metabolic machinery and are entirely dependent on hijacking the host's resources, including ATP, for replication.
Metabolic Target: The partitioning of host ATP between viral replication and essential host cell maintenance processes.
Experimental Protocol (Computational Modeling):
- Framework: A dynamic Flux Balance Analysis (FBA) framework was implemented within the COMETS (Computation of Microbial Ecosystems in Time and Space) platform to simulate the metabolic tension between a human host cell and SARS-CoV-2 [86].
- Model Setup: The genome-scale metabolic model of a human macrophage (iAB-AMØ-1410) was augmented with a detailed Virus Biomass Objective Function (VBOF) that included not only viral proteins and RNA but also, crucially, the lipid components of the virion [86].
- Objective Functions: The host cell's objective was defined as maximizing its ATP maintenance flux (a proxy for survival), while the virus's objective was to maximize the flux through the VBOF (a proxy for replication) [86].
- Dynamic Simulation: The model simulated the temporal dynamics of infection, tracking the consumption of nutrients (e.g., glucose, amino acids), the production of viral particles, and the depletion of intracellular ATP. The core innovation was a dynamic ATP partitioning parameter that controlled the degree to which the virus could hijack the host's ATP pool.
- Intervention Simulation: The model was used to in silico test the effect of inhibiting specific host metabolic pathways, such as lipid transport, on viral output and host cell survival.
Clinical Evidence and Quantitative Data: The computational model predicted that incorporating lipids into the viral biomass reaction increased the virion biomass production rate by 29.6% and altered the flux through 227 host metabolic reactions, highlighting the significant metabolic burden of viral replication [86]. The simulations revealed a critical threshold: if the viral demand for ATP drives the available ATP below the level required for host cell maintenance, the cell dies, abruptly terminating viral production. This model provides a mechanistic basis for exploring drugs that strategically push infected cells toward this threshold by exacerbating the virus-induced energy crisis.
Figure 2: ATP Competition Model in Viral Infection. This diagram depicts the metabolic competition for ATP between a host cell and an intracellular virus.
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents for Investigating Metabolism in Persistent Infections
| Reagent / Material | Function in Experimental Protocol | Specific Application Example |
|---|---|---|
| Gene Deletion Mutants (e.g., Δhyd) | To establish the essentiality of specific metabolic pathways for survival under dormancy-inducing conditions. | Validating the role of hydrogenase in microbial persistence via comparative viability studies [85]. |
| Luciferase-based ATP Assay Kits | To quantitatively measure intracellular ATP concentrations, providing a direct readout of the cellular energy state. | Monitoring ATP depletion in starved cultures or pathogen-infected host cells [86]. |
| Stable Isotope-Labeled Substrates (e.g., ¹³C-Glucose) | To trace the fate of nutrients through metabolic pathways using techniques like GC-MS or LC-MS, revealing flux distributions. | Determining if a pathogen switches carbon sources during dormancy or in response to an inhibitor. |
| Genome-Scale Metabolic Models (GEMs) | Computational frameworks to simulate and predict metabolic behavior under different genetic and environmental conditions. | Performing Flux Balance Analysis (FBA) to predict drug targets or simulate host-pathogen metabolic interactions [86] [87]. |
| Gas-tight Bioreactors | To maintain and precisely control the atmospheric composition (e.g., O₂, H₂, CO₂) during long-term microbial cultivation. | Studying the persistence of microbes on trace gases like atmospheric H₂ [85]. |
Discussion and Synthesis
The case studies presented herein underscore a common principle: successful persistence by pathogens is underpinned by metabolic flexibility and a finely tuned management of energy resources. The ability of bacteria to scavenge alternative energy sources like atmospheric H₂ and the fierce competition for ATP during viral infection are two manifestations of the same battle for metabolic supremacy.
Targeting these pathways offers a promising therapeutic strategy. In dormant bacteria, inhibiting maintenance metabolism (e.g., with small-molecule inhibitors of hydrogenases or other key energy-scavenging enzymes) could induce catabolic failure, driving the cells into a state of irreversible ATP depletion and death. Similarly, in viral infections, interventions that exacerbate the virus-induced energy drain could push infected cells to die prematurely, aborting the viral replication cycle. These approaches, while promising, require sophisticated tools for monitoring metabolic fluxes and intracellular energetics in real-time within complex host environments.
Clinical evidence from both bacteriology and virology confirms that the metabolic pathways supporting persistent infections are viable therapeutic targets. The emerging paradigm is to shift from targeting growth to targeting persistence itself, focusing on the unique metabolic vulnerabilities that emerge in dormant pathogens. Future research must focus on translating these findings from model systems to human infections, leveraging advanced metabolomics and computational modeling to design precise interventions that disrupt the energetic balance of persistent pathogens, ultimately leading to their eradication.
Bacterial persisters represent a transiently antibiotic-tolerant subpopulation that significantly contributes to chronic and relapsing infections, posing a major challenge in clinical treatments [12]. Unlike genetic resistance, persistence operates through phenotypic dormancy mechanisms, with ATP depletion and metabolic rewiring serving as central mediators of this tolerant state [8] [88]. Research has established that reduced intracellular ATP levels trigger a shift to a dormant-like state that enhances antibiotic tolerance, as persister cells maintain lower ATP levels than metabolically active vegetative cells [8] [89]. The complex relationship between bacterial metabolism and persistence necessitates sophisticated analytical approaches to unravel the mechanisms underlying this phenotype.
The integration of multi-omics data—encompassing genomics, transcriptomics, proteomics, and metabolomics—provides unprecedented opportunities to systematically decode the molecular networks governing persister formation and survival [90] [91]. Where single-omics approaches struggle to capture the complex interactions between different biological layers, multi-omics integration can reveal the sophisticated metabolic rewiring that occurs during the transition to persistence [90] [88]. This technical guide explores cutting-edge methodologies for integrating multi-omics data to predict and target bacterial persisters, with particular emphasis on ATP depletion and energy metabolism as central themes in persistent cell formation.
Theoretical Foundation: Metabolic Mechanisms of Persistence
ATP Depletion as a Central Driver of Persister Formation
Adenosine triphosphate (ATP) serves as a crucial indicator of cellular metabolic activity, and its depletion has been strongly associated with persister cell formation [8] [89]. Quercetin, a naturally occurring flavonoid, exemplifies this phenomenon by demonstrating a dose-dependent decrease in intracellular ATP levels in Staphylococcus aureus—reducing ATP by 22% and 36% at concentrations of 1 mM and 10 mM, respectively [8]. This ATP depletion correlates directly with increased persister cell formation across multiple antibiotic classes, with one study reporting 217-fold increased persistence in tobramycin-treated groups when combined with quercetin [8].
The molecular basis for quercetin-induced ATP depletion appears to involve disruption of electron transport chain function, impairment of oxidative phosphorylation, and interaction with ATP synthase [8]. Additional mechanisms include interference with the tricarboxylic acid (TCA) cycle and generation of reactive oxygen species that damage cellular components essential for energy metabolism [8]. This metabolic stress response induces a dormant state that limits energy-dependent processes required for antibiotic-mediated killing, including active drug uptake mechanisms that rely on proton motive force [8].
Metabolic Heterogeneity and Persister Subtypes
Persister cells exhibit significant metabolic heterogeneity, leading to their classification into distinct subtypes based on formation mechanisms and metabolic characteristics:
Table 1: Classification of Bacterial Persister Subtypes
| Persister Type | Formation Trigger | Metabolic State | Key Characteristics |
|---|---|---|---|
| Type I | Stationary phase entry; environmental stress | Non-growing or metabolically stagnant | Pre-formed during stationary phase; reversible dormancy [12] [7] |
| Type II | Stochastic processes during exponential growth | Slow-growing with reduced metabolism | Continuously generated throughout growth phase [12] [7] |
| Type III | Antibiotic-specific stress signals | Variable metabolic states | Specialized mechanisms for specific antibiotics; not necessarily slow-growing [7] |
This metabolic heterogeneity extends to energy generation pathways, with research indicating that while persisters exhibit reduced metabolic rates compared to exponentially growing cells, their survival still relies on maintained energy metabolism through the TCA cycle, electron transport chain, and ATP synthase activity [88]. The Crp/cAMP global regulatory system in Escherichia coli redirects persister cell metabolism from anabolism to oxidative phosphorylation, maintaining essential energy generation while downregulating biosynthetic pathways [88].
Signaling Networks and Metabolic Regulation in Persistence
Several key signaling pathways and molecular networks regulate the metabolic transitions into persistence:
Figure 1: Signaling Pathways Converging on Metabolic Persistence
The alarmone ppGpp serves as a critical mediator between metabolic state and persistence, activating toxin-antitoxin (TA) systems and other regulatory factors that drive metabolic downturn [89] [12]. Under nutrient limitation, cAMP levels increase through activation of adenylate cyclase (CyaA), forming complexes with the Crp receptor protein that activate catabolic genes supporting cellular energy maintenance [88]. Host-produced reactive oxygen and nitrogen species (ROS/RNS) further induce metabolic collapse and antibiotic tolerance across numerous bacterial species, including S. aureus, Mycobacterium tuberculosis, and Salmonella enterica Typhimurium [92].
Multi-Omics Integration Frameworks for Persister Research
Computational Strategies for Multi-Omics Data Integration
The complexity of persister metabolism requires advanced computational approaches that can integrate diverse omics datasets to reconstruct the molecular networks underlying persistence:
Table 2: Multi-Omics Integration Strategies for Persister Research
| Integration Approach | Methodology | Applications in Persistence Research | Tools/Platforms |
|---|---|---|---|
| Data-based Integration | Concatenates different omics datasets into a unified matrix | Identifies cross-omics signatures of ATP depletion; reveals coordinated metabolic changes | MEMINEX [91] |
| Model-based Integration | Combines individual models for each omics dataset | Models metabolic flux during persistence transitions; predicts regulatory networks | OBaNK [91] |
| Network-based Methods | Constructs interaction networks between heterogeneous omics types | Maps TA system interactions; identifies key metabolic bottlenecks | Bayesian Networks [91] |
| Correlation/Association | Elucidates similarity across omics types via group membership | Correlates transcriptomic and proteomic profiles with ATP levels | OmicsNet [91] |
| Dimension Reduction | Discovers latent features combining different omics types | Characterizes persister subpopulations based on metabolic states | Cox-nnet, DeepProg [93] |
Probabilistic graph representations coupled with external knowledge bases offer particularly powerful approaches for multi-omics integration in persister research [91]. The MEMINEX algorithm exemplifies this approach by using Bayesian networks to integrate transcriptomic, proteomic, and metabolomic data while incorporating prior knowledge about molecular interactions [91]. This method has successfully identified active toxin/antitoxin systems and metabolic pathway interactions in bacterial communities under stress conditions [91].
Experimental Workflows for Multi-Omics in Persistence Studies
A standardized workflow for generating and integrating multi-omics data in persister research ensures reproducibility and cross-study comparability:
Figure 2: Multi-Omics Experimental Workflow
This workflow begins with careful persister induction using well-characterized stressors such as quercetin for ATP depletion [8], antibiotic exposure [12], or host-mimicking conditions including ROS/RNS generation [92]. Parallel sampling for different omics analyses is critical, as technical variation can obscure true biological signals in integrated analyses [91]. For transcriptomics, RNA isolation followed by high-throughput sequencing captures gene expression patterns, while proteomics employs mass spectrometry to quantify protein abundance, and metabolomics utilizes mass spectrometry or NMR to profile metabolic states [91].
Single-Cell and Spatial Multi-Omics Technologies
Bulk omics approaches average signals across cell populations, potentially obscuring the inherent heterogeneity of persister subpopulations [90]. Single-cell multi-omics technologies overcome this limitation by enabling simultaneous measurement of transcriptomic, epigenomic, and proteomic information from individual cells [90]. This approach is particularly valuable for characterizing the metabolic diversity within persister populations and identifying rare deep-persister states [12].
Spatial multi-omics further extends these capabilities by preserving the anatomical context of bacterial populations within host tissues or biofilms [90]. Spatial transcriptomics and proteomics technologies map molecular profiles to specific tissue locations, revealing how microenvironmental niches influence persister formation through local variations in nutrient availability, immune cell proximity, and antibiotic penetration [90]. These technologies are especially relevant for studying chronic biofilm-associated infections where spatial organization significantly impacts bacterial metabolism and antibiotic tolerance [12].
Analytical Methodologies for Persister Metabolism
Metabolomic and Isotopolog Profiling Techniques
Metabolomic approaches provide the most direct window into the physiological state of persister cells, with 13C-isotopolog profiling offering particularly powerful insights into metabolic pathway activities [89]. This technique involves feeding 13C-labeled carbohydrates to bacterial cultures and analyzing the labeling patterns of intermediate metabolites to deduce relative metabolic fluxes [89]. Application of this method to stationary-phase S. aureus cells challenged with daptomycin revealed active amino acid anabolism, glycolysis, TCA cycle, and pentose phosphate pathway activity despite the non-growing state [89].
Isotopolog profiling has demonstrated that persister cells maintain active energy metabolism with specific pathway alterations. Analysis of Asp and Glu labeling patterns indicated increased TCA cycle activity in S. aureus persisters, suggesting metabolic rewiring rather than complete metabolic shutdown [89]. Similar approaches in E. coli have elucidated the role of the Crp/cAMP complex in redirecting metabolism toward oxidative phosphorylation during persistence [88].
Proteogenomic Integration for Persister Marker Discovery
Proteogenomic approaches integrate genomic and proteomic data to discover novel protein coding regions and validate gene models, offering unique insights into persister-specific molecular mechanisms [90]. This methodology is particularly valuable for identifying truncated proteins, sequence variants, and post-translational modifications that may influence metabolic activity in persister cells.
In practice, proteogenomic analysis involves custom database construction from genomic and transcriptomic sequences, followed by mass spectrometry-based peptide identification and statistical validation [90]. Applied to persister research, this approach can reveal persister-specific protein isoforms resulting from alternative translation initiation, programmed frameshifts, or proteolytic processing that modulate metabolic enzyme activity and contribute to ATP depletion.
Network Analysis of Multi-Omics Data
Network-based analysis of multi-omics data reconstructs the complex interaction webs connecting different molecular layers in persister cells [91]. Bayesian networks are particularly well-suited for this task as they can capture both linear and nonlinear interactions in stochastic systems obscured by biological noise [91]. The MEMINEX algorithm implements this approach by using probabilistic graph representations and external knowledge to deduce multifarious interaction networks from multi-omics data [91].
Key applications of network analysis in persister research include:
- Identifying master regulatory nodes that control metabolic transitions into persistence
- Mapping cross-talk between TA systems and metabolic pathways
- Revealing bottlenecks in energy generation pathways during ATP depletion
- Predicting network fragility points that could be targeted for anti-persister therapies
Experimental Protocols for Key Investigations
Protocol 1: Assessing ATP Dynamics in Bacterial Persisters
Objective: Quantify intracellular ATP levels during persister formation and identify correlations with antibiotic tolerance.
Materials and Reagents:
- Bacterial strains (e.g., Staphylococcus aureus JE2)
- Quercetin (1-10 mM in DMSO) as ATP-depleting agent [8]
- ATP determination kit (luciferase-based assay)
- Antibiotics for persister selection (oxacillin, ciprofloxacin, tobramycin) [8]
- Bioluminescent reporter strains (e.g., JE2-lux) for metabolic activity monitoring [92]
Methodology:
- Culture bacteria to mid-exponential phase in appropriate medium
- Treat with quercetin (0-10 mM) for 2-4 hours to induce ATP depletion [8]
- Measure intracellular ATP levels using luciferase-based assay
- Parallelly assess persister frequencies by antibiotic challenge (5-100× MIC for 24h)
- Correlate ATP depletion levels with persister formation across conditions
- Validate using bioluminescent reporters to monitor metabolic activity in real-time [92]
Data Analysis: Calculate fold-change in persister frequency relative to ATP depletion. Use linear regression to model relationship between ATP levels and antibiotic tolerance.
Protocol 2: Multi-Omics Profiling of Persister Metabolic States
Objective: Generate integrated transcriptomic, proteomic, and metabolomic profiles of persister cells to map metabolic rewiring.
Materials and Reagents:
- RNAlater stabilization solution for transcriptomics
- Lysis buffer with protease inhibitors for proteomics
- Cold methanol:water (4:1) for metabolomic quenching
- 13C-labeled glucose or other carbon sources for isotopolog profiling [89]
- LC-MS/MS systems for proteomic and metabolomic analysis
- RNA sequencing library preparation kits
Methodology:
- Induce persister formation through antibiotic exposure (e.g., 5× MIC ampicillin for 4h) or metabolic stressors (e.g., quercetin) [8]
- Separate persisters from non-persisters using density gradient centrifugation or fluorescence-activated cell sorting
- Collect triplicate samples for each omics platform:
- Transcriptomics: Extract RNA, prepare sequencing libraries, perform RNA-seq
- Proteomics: Digest proteins, desalt peptides, perform LC-MS/MS
- Metabolomics: Extract metabolites, perform LC-MS analysis in positive and negative ion modes
- For flux analysis, incorporate 13C-labeled substrates during persister formation [89]
Data Integration:
- Apply batch correction across datasets
- Perform multivariate statistical analysis (PCA, OPLS-DA)
- Integrate using network-based approaches (Bayesian networks) [91]
- Map changes to metabolic pathways (TCA cycle, electron transport chain, ATP synthesis)
Protocol 3: High-Throughput Screening for Persister-Modulating Compounds
Objective: Identify compounds that alter persister metabolism and sensitize them to antibiotics.
Materials and Reagents:
- Bioluminescent bacterial reporter strains (e.g., JE2-lux) [92]
- 384-well assay plates
- Compound libraries (≥4,700 drug-like compounds) [92]
- Cell viability assay kits (e.g., resazurin-based)
- Antibiotics with different mechanisms of action
Methodology:
- Culture bioluminescent reporter bacteria to mid-exponential phase
- Infect mammalian macrophages (for intracellular persister models) or seed directly (for planktonic models)
- Add compound libraries (1-10 μM) with and without sub-MIC antibiotics
- Measure bioluminescence (metabolic activity) and viability at 4h and 24h [92]
- Counter-screen for host cytotoxicity using viability assays
- Validate hits in secondary assays with clinical isolates
Hit Selection Criteria:
- ≥1.5-fold increase in bacterial metabolic activity without cytotoxicity [92]
- ≥10-fold enhancement of antibiotic killing without promoting bacterial outgrowth alone
- Activity against multiple bacterial strains and persister subtypes
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Persister and Metabolic Studies
| Reagent/Category | Specific Examples | Research Application | Key References |
|---|---|---|---|
| ATP-Depleting Compounds | Quercetin (1-10 mM), Sodium arsenate (0-0.5 mM) | Induce metabolic dormancy and persister formation | [8] [92] |
| Metabolic Reporters | JE2-lux bioluminescent strain, ATP-sensitive luciferase assays | Monitor bacterial metabolic activity and energy status in real-time | [92] |
| Persistence Inducers | Ampicillin (5-100× MIC), Ciprofloxacin, Tobramycin | Select for and study antibiotic-tolerant persister subpopulations | [8] [12] |
| Integration Algorithms | MEMINEX, OBaNK, Bayesian Networks | Integrate multi-omics datasets and reconstruct molecular interaction networks | [91] |
| 13C-Labeled Substrates | 13C-glucose, 13C-acetate, 13C-amino acids | Trace metabolic fluxes and pathway activities through isotopolog profiling | [89] |
| Host-Pathogen Models | Bone marrow-derived macrophages, murine infection models | Study intracellular persisters in host-mimicking environments | [92] |
Future Perspectives and Concluding Remarks
The integration of multi-omics data represents a paradigm shift in persister research, moving beyond simplistic dormancy models to reveal the sophisticated metabolic rewiring that enables bacterial survival under antibiotic pressure [88]. Future research directions should prioritize several key areas:
First, temporal multi-omics profiling capturing the transition into and out of persistence will reveal dynamic metabolic checkpoints that could be targeted therapeutically. Second, single-cell multi-omics approaches will elucidate the metabolic heterogeneity within persister populations, potentially identifying specialized metabolic programs in deep-persister states [90]. Third, host-pathogen integration of multi-omics data will illuminate how host-derived stresses (ROS/RNS, nutrient limitation) shape bacterial metabolic networks during infection [92].
From a therapeutic perspective, compounds like KL1 that modulate host responses to resuscitate intracellular bacteria represent promising adjuvant strategies [92]. Similarly, targeting the Crp/cAMP regulatory system or specific TCA cycle enzymes may disrupt energy metabolism essential for persister survival [88]. The continuing development of computational integration methods, particularly probabilistic graph models incorporating external knowledge, will be essential for translating multi-omics data into actionable insights for persistent infection management [91].
In conclusion, the strategic integration of multi-omics data provides an unprecedented window into the metabolic adaptations underlying bacterial persistence. By leveraging these powerful approaches, researchers can systematically map the molecular networks controlling ATP depletion and energy metabolism in persister cells, ultimately enabling predictive targeting of this elusive bacterial subpopulation to improve treatment outcomes for persistent infections.
Conclusion
The investigation of ATP depletion and energy metabolism represents a paradigm shift in understanding and combating bacterial persistence. Key takeaways reveal that dormancy is an active metabolic state regulated by precise molecular mechanisms, including ATP-mediated stringent response and protein aggregation, rather than simple inactivity. Therapeutically, targeting bacterial energetics offers promising avenues, notably through metabolite-driven resuscitation that re-sensitizes persisters to conventional antibiotics. Future research must prioritize translating these mechanistic insights into clinical strategies that overcome biofilm barriers and physiological heterogeneity. Integrating metabolic approaches with traditional antibiotics presents the most viable path toward eradicating persistent infections, ultimately addressing a critical bottleneck in infectious disease management and antimicrobial stewardship.
References
- 1. : molecular mechanisms and therapeutic... Bacterial persisters [https://www.nature.com/articles/s41392-024-01866-5?error=cookies_not_supported&code=4bbdd9bc-caa6-4305-9e89-2929af9a7100]
- 2. Integrative biology of persister cell formation - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC9470271/]
- 3. Slow and steady wins the race: an examination of bacterial persistence [https://www.aimspress.com/microbiology/article/1345/fulltext.html]
- 4. Assessing the relative importance of bacterial , persistence... resistance [https://pmc.ncbi.nlm.nih.gov/articles/PMC9653239/]
- 5. Are Bacterial Persisters Dormant Cells Only? [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.708580/full]
- 6. Molecular mechanism and application of emerging technologies in... [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/s12866-024-03628-3]
- 7. From Dormancy to Eradication: Strategies for Controlling ... [https://www.mdpi.com/2304-8158/14/6/1075]
- 8. Metabolic Stress Induced by Quercetin Enhances Dormancy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12108273/]
- 9. Observation of persister cell histories reveals diverse ... [https://elifesciences.org/articles/79517]
- 10. Metabolite-driven reprogramming of bacterial persisters [https://www.sciencedirect.com/science/article/abs/pii/S1368764625001256]
- 11. Bioenergetic stress potentiates antimicrobial resistance ... [https://www.nature.com/articles/s41467-025-60302-6]
- 12. Bacterial persisters: molecular mechanisms and ... [https://www.nature.com/articles/s41392-024-01866-5]
- 13. stress symptoms and its related responses induced by (over ... [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-024-02370-9]
- 14. Molecular Mechanisms of Bacterial Clinical Pathogens ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8350155/]
- 15. Composition and liquid-to-solid maturation of protein ... [https://www.nature.com/articles/s41467-025-56387-8]
- 16. Boosting energy metabolism and biosynthesis in diverse ... [https://www.nature.com/articles/s41467-025-62638-5]
- 17. Common pathogenic bacteria-induced reprogramming of the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10689525/]
- 18. Hallmarks of Metabolic Reprogramming and Their Role in ... [https://www.mdpi.com/1999-4915/14/3/602]
- 19. Spatial quantitative metabolomics enables identification of ... [https://www.nature.com/articles/s42255-025-01340-8]
- 20. Phenotypic heterogeneity in persisters: a novel 'hunker' ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8767447/]
- 21. New study uncovers mechanisms of bacterial dormancy ... [https://press.vib.be/new-study-uncovers-mechanisms-of-bacterial-dormancy-involving-protein-aggregation]
- 22. Phenotypic heterogeneity in the bacterial oxidative stress ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10935545/]
- 23. Quantitation and Comparison of Phenotypic Heterogeneity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6933826/]
- 24. Bacteriophage-driven microbial phenotypic heterogeneity [https://www.nature.com/articles/s41522-025-00727-5]
- 25. Phenotypic heterogeneity follows a growth-viability tradeoff ... [https://www.nature.com/articles/s41467-024-50602-8]
- 26. The Thinking Undead: How Dormant Bacteria Calculate Their ... [https://today.ucsd.edu/story/the-thinking-undead-how-dormant-bacteria-calculate-their-return-to-life]
- 27. Unexpected Activity in 'Dead' Bacteria Detected by Scientists [https://www.sciencealert.com/unexpected-activity-in-dead-bacteria-detected-by-scientists]
- 28. ATP Is a Major Determinant of Phototrophic Bacterial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10128053/]
- 29. ATP, ADP and AMP profiling for diagnostic applications [https://www.sciencedirect.com/science/article/pii/S0165993625003929]
- 30. A Novel HPLC-MS/MS Method for the Intracellular ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12285933/]
- 31. ATP biosensor reveals microbial energetic dynamics and ... [https://www.nature.com/articles/s41467-024-49579-1]
- 32. Structural basis for bacterial energy extraction from ... [https://www.nature.com/articles/s41586-023-05781-7]
- 33. Challenges and Advances for Genetic Engineering of Non ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5662904/]
- 34. Synthetic Biology-Based Approaches to Investigate Host– ... [https://www.mdpi.com/2674-0583/3/1/4]
- 35. Synthetic biology-inspired cell engineering in diagnosis ... [https://www.nature.com/articles/s41392-023-01375-x]
- 36. Quantitative metabolic fluxes regulated by trans-omic networks [https://pmc.ncbi.nlm.nih.gov/articles/PMC9022981/]
- 37. Interpreting Expression Data with Metabolic Flux Models [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1000489]
- 38. Backtracking metabolic dynamics in single cells predicts ... [https://www.nature.com/articles/s41467-025-64225-0]
- 39. Mini review: Persister cell control strategies [https://www.frontiersin.org/journals/pharmacology/articles/10.3389/fphar.2025.1706115/full]
- 40. Bacterial Metabolic Heterogeneity: Origins and Applications in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7485380/]
- 41. Antibiotic persisters are metabolically active, non-growing ... [https://www.sciencedirect.com/science/article/pii/S0924857924003029]
- 42. Integrative single-cell metabolomics and phenotypic ... [https://www.nature.com/articles/s41467-025-57992-3]
- 43. Bacterial single-cell RNA sequencing captures biofilm ... [https://www.nature.com/articles/s41467-024-54581-8]
- 44. Bacterial Metabolism and Antibiotic Efficacy - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC6990394/]
- 45. Host Cell Oxidative Stress Induces Dormant Staphylococcus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8865412/]
- 46. ATP-Dependent Dynamic Protein Aggregation Regulates ... [https://www.sciencedirect.com/science/article/pii/S1097276518308827]
- 47. Metabolite-Enabled Eradication of Bacterial Persisters by ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3145328/]
- 48. Unveiling the critical roles of cellular metabolism ... [https://www.nature.com/articles/s44259-024-00034-7]
- 49. Visualization of ATP levels inside single living cells with ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2735558/]
- 50. Visualization of intracellular ATP dynamics in different ... [https://www.sciencedirect.com/science/article/pii/S0085253824004095]
- 51. Recent Advances in Bacterial Persistence Mechanisms - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC10531727/]
- 52. Role of protein aggregates in bacteria [https://pubmed.ncbi.nlm.nih.gov/40324851/]
- 53. Role of protein aggregates in bacteria [https://www.sciencedirect.com/science/article/abs/pii/S1876162324000993]
- 54. What are the different methods of determining protein ... [https://www.enzo.com/note/what-are-the-different-methods-of-determining-protein-aggregation/]
- 55. Measuring Protein Aggregation and Stability Using High- ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9149252/]
- 56. Massive experimental quantification allows interpretable ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12042874/]
- 57. Quantitative analysis of persister fractions suggests different ... [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/1471-2180-13-25]
- 58. Targeting bacterial membrane function: an underexploited mechanism for treating persistent infections - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3496266/]
- 59. Phage Paride can kill dormant, antibiotic-tolerant cells of ... [https://www.nature.com/articles/s41467-023-44157-3]
- 60. Revived Amplicon Sequence Variants Monitoring in Closed ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10055844/]
- 61. Release of extracellular ATP by bacteria during growth | BMC Microbiology | Full Text [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/1471-2180-13-301]
- 62. Antibiotics May Help Bacteria Accelerate Treatment ... [https://www.insideprecisionmedicine.com/topics/patient-care/antibiotics-may-help-bacteria-accelerate-treatment-resistance/]
- 63. Mitigating ROS signalling pathway-mediated defence ... [https://link.springer.com/article/10.1007/s11101-025-10179-6]
- 64. Ferroptosis-new avenues for the treatment of multidrug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12537265/]
- 65. Treatment strategies targeting persister cell formation in ... [https://pubmed.ncbi.nlm.nih.gov/33022189/]
- 66. A Strategy to Suppress Staphylococcus aureus Persister ... [https://www.mdpi.com/1422-0067/26/13/6335]
- 67. Environmental conditions define the energetics of bacterial ... [https://www.sciencedirect.com/science/article/pii/S0006349523004113]
- 68. Bioavailability Enhancement Techniques for Poorly Aqueous ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9495787/]
- 69. What are the methods used for enhancement of ... [https://synapse.patsnap.com/article/what-are-the-methods-used-for-enhancement-of-bioavailability]
- 70. Carbon and energy metabolism of atp mutants ... [https://pubmed.ncbi.nlm.nih.gov/1447134/]
- 71. The msaABCR Operon Regulates Persister Formation by ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.657753/full]
- 72. Metabolic Stress Induced by Quercetin Enhances ... [https://www.mdpi.com/2079-6382/14/5/424]
- 73. Energy Metabolism and Drug Efflux in Mycobacterium ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3993223/]
- 74. ATPase mediates regulation of fermentation and energy ... [https://www.nature.com/articles/s41598-025-21047-w]
- 75. Defects in energy metabolism increase the susceptibility of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12502701/]
- 76. Environmental conditions define the energetics of bacterial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10465703/]
- 77. Representation of Dormant and Active Microbial Dynamics for ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0089252]
- 78. Phenotypic screening of compound libraries as a platform ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10942738/]
- 79. Discovery of antibiotics that selectively kill metabolically ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11031330/]
- 80. Screening an Established Natural Product Library Identifies ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8330956/]
- 81. Prenylated flavonoids combat metabolically dormant ... [https://the-innovation.org/article/doi/10.59717/j.xinn-life.2025.100154]
- 82. Intracellular ATP concentration is a key regulator of bacterial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11656805/]
- 83. Microbiological Applications: Bacterial Metabolism [https://www.bmglabtech.com/en/blog/microbiological-applications-for-bacterial-metabolism-on-a-microplate-reader/]
- 84. The Basics: In Vitro Translation [https://www.thermofisher.com/us/en/home/references/ambion-tech-support/large-scale-transcription/general-articles/the-basics-in-vitro-translation.html]
- 85. Uncovering the Metabolic Strategies of the Dormant Microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6529542/]
- 86. Dynamic metabolic modeling of ATP allocation during viral infection - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC11601281/]
- 87. Computational Approaches for Understanding Energy ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3906216/]
- 88. Unraveling CRP/cAMP-mediated metabolic regulation in ... [https://elifesciences.org/articles/99735]
- 89. Metabolic aspects of bacterial persisters - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4205924/]
- 90. Advances in Integrated Multi-omics Analysis for Drug-Target ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11201992/]
- 91. Integration of Meta-Multi-Omics Data Using Probabilistic ... [https://www.mdpi.com/2073-4409/12/15/1998]
- 92. A host-directed adjuvant sensitizes intracellular bacterial ... [https://www.nature.com/articles/s41564-025-02124-2]
- 93. Strategies to integrate multi-omics data for patient survival ... [https://arxiv.org/abs/2008.12455]