Matrix Guardians: Decoding the Structural and Protective Roles of Alginate, Psl, and Pel in Pseudomonas aeruginosa Biofilms
This article provides a comprehensive analysis of the three key exopolysaccharides—alginate, Psl, and Pel—that constitute the biofilm matrix of the opportunistic pathogen Pseudomonas aeruginosa.
Matrix Guardians: Decoding the Structural and Protective Roles of Alginate, Psl, and Pel in Pseudomonas aeruginosa Biofilms
Abstract
This article provides a comprehensive analysis of the three key exopolysaccharides—alginate, Psl, and Pel—that constitute the biofilm matrix of the opportunistic pathogen Pseudomonas aeruginosa. Tailored for researchers and drug development professionals, we dissect the unique and synergistic functions of these polymers, from foundational biology and structural roles to their critical contribution in antibiotic tolerance and resistance to host defenses. The scope extends to current methodological approaches for studying these components, the challenges in targeting them, and the promising validation of novel therapeutic strategies that exploit these matrix polymers as antibiotic adjuvants. Understanding this complex exopolysaccharide landscape is paramount for developing next-generation treatments against persistent biofilm-mediated infections, particularly in cystic fibrosis.
The EPS Trinity: Unveiling the Structure, Biosynthesis, and Core Functions of Alginate, Psl, and Pel
Within the realm of infectious disease research, the biofilm-forming opportunistic pathogen Pseudomonas aeruginosa presents a significant challenge, particularly in the context of cystic fibrosis (CF) and nosocomial infections. The resilience of this bacterium is largely attributed to its self-produced extracellular matrix, a critical component of which is a trio of exopolysaccharides: alginate, Psl, and Pel [1] [2]. These polymers form the structural backbone of biofilms, conferring enhanced tolerance to antibiotics and host immune defenses [3]. A precise understanding of their unique chemical compositions, architectural roles, and biosynthetic pathways is not merely an academic exercise but a fundamental prerequisite for developing targeted therapeutic strategies to disrupt these protective communities [2]. This review synthesizes current knowledge to explicitly define the chemical identity of each polysaccharide and delineate their distinct contributions to the biofilm matrix, providing a framework for ongoing research and therapeutic development.
Chemical and Structural Profiles of P. aeruginosa Exopolysaccharides
The biofilm matrix of P. aeruginosa is primarily composed of three exopolysaccharides—alginate, Psl, and Pel—each with a distinct chemical structure that dictates its specific functional role within the microbial community. The defining characteristics of these polymers are summarized in Table 1.
Table 1: Comparative Chemical and Functional Profiles of P. aeruginosa Exopolysaccharides
| Feature | Alginate | Psl | Pel |
|---|---|---|---|
| Chemical Composition | Non-repetitive, linear copolymer of β-1,4-linked D-mannuronic acid (M) and its C5 epimer, L-guluronic acid (G) [1] [4]. | Branched pentasaccharide repeating unit composed of D-mannose, D-glucose, and L-rhamnose [2] [4]. | Linear homopolymer of partially de-N-acetylated α-1,4-linked N-acetylgalactosamine (GalNAc) [5]. |
| Primary Monosaccharides | D-mannuronic acid, L-guluronic acid [1]. | D-mannose, D-glucose, L-rhamnose [2]. | Galactosamine (GalN) and N-acetylgalactosamine (GalNAc) in a ~2:1 ratio [5]. |
| Charge Property | Anionic (polyanionic) [6]. | Neutral [6]. | Cationic at physiological pH [2] [5]. |
| Key Genetic Loci | alg gene cluster (e.g., algD, alg8) [7] [8]. |
psl operon (pslA-O) [1] [2]. |
pel operon (pelA-G) [1] [9]. |
| Primary Structural Role | Forms a hydrogel-like matrix that protects mucoid strains; contributes to biofilm viscosity and mechanical stability [1] [6]. | Acts as a primary scaffold for cell-cell and cell-surface interactions; crucial for initial attachment and microcolony formation in non-mucoid strains like PAO1 [1] [8]. | Serves as a structural scaffold in strains like PA14; crucial for pellicle formation and provides structural integrity through ionic cross-linking with eDNA [2] [3]. |
Alginate: The Anionic Protector
Alginate is a high molecular weight, linear copolymer. Its non-repetitive structure and the degree of O-acetylation on the mannuronic acid residues contribute to its physical properties [1] [4]. This polysaccharide is most notably overproduced by mucoid strains of P. aeruginosa, which frequently emerge in the lungs of CF patients [1]. The overproduction, often resulting from mutations in the negative regulator mucA, correlates with a worsened clinical prognosis [1]. Alginate forms a viscous, gelatinous capsule around cells, functioning as a physical barrier that scavenges reactive oxygen species and impedes phagocytosis by immune cells [1] [6]. Its anionic nature and high water retention capacity allow it to form a hydrogel that significantly contributes to the mechanical stability and resistance of biofilms [6].
Psl: The Neutral Scaffold
Psl is a neutral, branched exopolysaccharide that plays a critical role in the biofilm development of non-mucoid strains such as PAO1 [1] [2]. Genetic and biochemical studies have shown that the 15-gene psl operon is essential for the synthesis of this mannose- and galactose-rich polysaccharide [1]. Psl is produced during planktonic growth and is displayed on the cell surface in a helical pattern, which facilitates the initial attachment to surfaces and the formation of microcolonies [2] [4]. As the biofilm matures, Psl functions as a structural scaffold, holding cells together within the matrix [1]. It is strategically localized to the caps of mushroom-shaped microcolonies in mature biofilms, forming a peripheral meshwork that is crucial for maintaining the three-dimensional architecture [7]. Furthermore, Psl can cross-link with the adhesin CdrA and extracellular DNA (eDNA), further reinforcing the biofilm structure [2].
Pel: The Cationic Cross-Linker
Pel is a cationic, linear polysaccharide composed of a dimeric repeat of α-1,4 linked galactosamine and N-acetylgalactosamine [5]. This composition was definitively established through advanced isolation techniques and glycoside hydrolase digestion, overcoming previous challenges related to its insolubility [5]. The positive charge of Pel, resulting from the partial de-N-acetylation of the GalNAc subunits, is a key functional attribute [2] [5]. This charge enables Pel to engage in ionic cross-linking with other negatively charged matrix components, most notably eDNA, which is critical for the structural integrity of the biofilm in strains like PA14 [2] [3]. Pel is essential for pellicle formation at the air-liquid interface and, in certain strains, serves as the primary structural polysaccharide [1] [3]. Its ability to bind and protect eDNA from degradation by nucleases also contributes to the stability of the biofilm matrix and enhances tolerance to aminoglycoside antibiotics [2] [3].
Functional Architecture and Synergistic Roles in Biofilms
The exopolysaccharides of P. aeruginosa do not function in isolation; their combined actions and strain-specific expression create a robust and adaptive biofilm architecture. The physical and functional interactions between these components are critical for the emergent properties of the biofilm community.
Table 2: Functional Redundancy and Synergy in Polysaccharide-Deficient Mutants
| Genotype | Biofilm Phenotype | Key Compensatory Mechanism | Experimental Evidence |
|---|---|---|---|
| PAO1 ΔpslA | Severe defect in initial attachment and mature biofilm structure; reduced biomass and thickness [7] [8]. | Enhanced production of Pel polysaccharide [7]. | Transcriptional profiling and phenotypic analysis of biofilm variants [7]. |
| PAO1 ΔpelF | Moderate reduction in biofilm biomass, but architecture is largely maintained [8] [3]. | Psl serves as the primary structural scaffold; functional redundancy is minimal in this strain [3]. | Crystal violet staining and microscopic analysis show Psl is the dominant matrix component in PAO1 [8] [3]. |
| PAO1 Δalg8 | No significant difference in biofilm formation compared to wild-type under standard conditions [7] [8]. | Psl and Pel are sufficient for biofilm formation in non-mucoid strains [1] [7]. | Biofilm mass and architecture are unaffected in flow cell and microtiter dish assays [7] [8]. |
| PA14 ΔpelB | Severe biofilm deficiency; inability to form structured pellicles or mature biofilms [3]. | PA14 has a natural truncation in the psl operon and cannot produce Psl, leaving no redundant scaffold [3]. |
Optical tweezer assays show loss of cell-cell interactions; significant reduction in biofilm biomass [3]. |
The strain-dependent hierarchy of these polysaccharides is evident. In the laboratory strain PAO1, Psl acts as the dominant structural component, with Pel playing a more auxiliary role [8] [3]. Conversely, in strain PA14, which is naturally incapable of producing Psl, Pel is indispensable as the primary structural scaffold [3]. The regulation of their production is complex and interconnected. Evidence suggests an inverse regulatory relationship between alginate and Psl, as indicated by a 30% decrease in Psl promoter activity in an alginate-overproducing mucA mutant [7]. Furthermore, the absence of one polysaccharide can lead to the increased production of another, as seen in psl mutants that overproduce Pel [7]. This functional redundancy underscores the evolutionary advantage bestowed by maintaining multiple polysaccharide systems, ensuring biofilm integrity under fluctuating environmental conditions.
Experimental Methodologies for Isolation and Characterization
A comprehensive understanding of exopolysaccharide function is rooted in robust methodologies for their isolation, purification, and characterization. The following protocols represent key experimental approaches cited in the literature.
Protocol for Pel Isolation and Structural Analysis
The precise chemical structure of Pel was historically difficult to determine due to its insolubility. A modified isolation and analysis protocol was pivotal in elucidating its true composition [5].
- Bacterial Strain and Culture: Use an engineered Pel overexpression strain (e.g., PAO1 ΔwspF Δpsl PBADpel). Inoculate cultures and induce Pel expression with arabinose [5].
- Polysaccharide Isolation: Centrifuge cultures to remove cells. Subject the supernatant to extensive dialysis against deionized water to remove contaminating sugars from peptidoglycan fragments. Precipitate the polysaccharide from the dialysate using isopropanol [5].
- Solubilization and Digestion: Chemically re-N-acetylate the insoluble Pel polymer to enhance solubility. Treat the re-N-acetylated Pel with the recombinantly expressed glycoside hydrolase domain of PelA (PelAh) to hydrolyze it into short, soluble oligosaccharides [5].
- Structural Determination:
- Glycosyl Composition and Linkage Analysis: Use gas chromatography-mass spectrometry (GC-MS) following reductive amination, hydrolysis, re-N-acetylation, and derivatization. This identifies monosaccharides as GalN and GalNAc and confirms 1-4 linkages [5].
- Anomeric Configuration Analysis: Employ 1H Nuclear Magnetic Resonance (NMR) spectroscopy on the PelAh-generated oligosaccharides. Compare the spectrum and coupling constants to a synthetic α-1,4-linked GalNAc oligosaccharide standard to confirm the α-stereochemistry [5].
Protocol for Genetic Construction of Polysaccharide-Deficient Mutants
Generating isogenic mutants is fundamental for dissecting the individual and synergistic roles of each polysaccharide. A standard method involves gene replacement via homologous recombination [7].
- Mutant Construction:
- Amplify Flanking Regions: For the target gene (e.g.,
pelF,pslA,alg8), amplify ~1 kb DNA regions upstream (N) and downstream (C) by PCR. - Clone Flanking Regions: Ligate the N and C fragments into a suicide vector containing a gentamicin resistance cassette (
aacC1) and a sucrose-sensitivity gene (sacB). - Conjugate and Integrate: Transfer the resulting plasmid into P. aeruginosa via conjugation or electroporation. Select for single-crossover integrants on gentamicin-containing plates.
- Counter-Selection: Plate integrants on mineral salt medium with sucrose to select for a second crossover event and the loss of the vector backbone. Verify gene replacement by PCR [7].
- Amplify Flanking Regions: For the target gene (e.g.,
- Generation of Multiple Mutants: To create double or triple mutants, repeat the process sequentially in existing markerless single mutants, using a Flp recombinase system to excise the antibiotic resistance cassette after each round of mutagenesis [7].
Experimental Workflow for Functional Biofilm Analysis
The following diagram outlines a standard integrated workflow for analyzing the role of exopolysaccharides in biofilm formation, from genetic manipulation to phenotypic and structural characterization.
The Scientist's Toolkit: Key Research Reagents and Materials
Advancing research on P. aeruginosa exopolysaccharides relies on a specific toolkit of genetic constructs, enzymatic tools, and analytical reagents.
Table 3: Essential Research Reagents for Exopolysaccharide Studies
| Reagent / Material | Function and Application | Specific Examples / Notes |
|---|---|---|
| Engineered Bacterial Strains | Used to overexpress or delete specific polysaccharides for functional studies. | PAO1 ΔwspF Δpsl PBADpel (for inducible Pel expression) [5]; PAO1/PA14 isogenic single, double, and triple mutants (ΔpelF, ΔpslA, Δalg8) [7] [8]. |
| Glycoside Hydrolases | Enzyme-based degradation of specific polysaccharides for structural analysis or biofilm disruption. | Alginate lyase (degrades alginate) [1]; PelAh (the hydrolase domain of PelA, cleaves Pel for structural analysis) [5]. |
| Lectins and Fluorescent Probes | Visualization and localization of polysaccharides within biofilms using microscopy. | Wisteria floribunda (WFL) lectin (binds terminal GalNAc, specific for Pel) [5]; Antibodies or other lectins for Psl staining [1]. |
| Analytical Standards | Reference materials for validating chemical structures. | Synthetic α-1,4-linked N-acetylgalactosamine (GalNAc) oligosaccharides (for NMR comparison with Pel) [5]. |
| Specialized Growth Systems | Cultivation of biofilms under controlled, reproducible conditions for architectural and mechanical testing. | Flow-cell reactors for CLSM and microrheology [6]; Microtiter dishes for static biofilm assays [3]. |
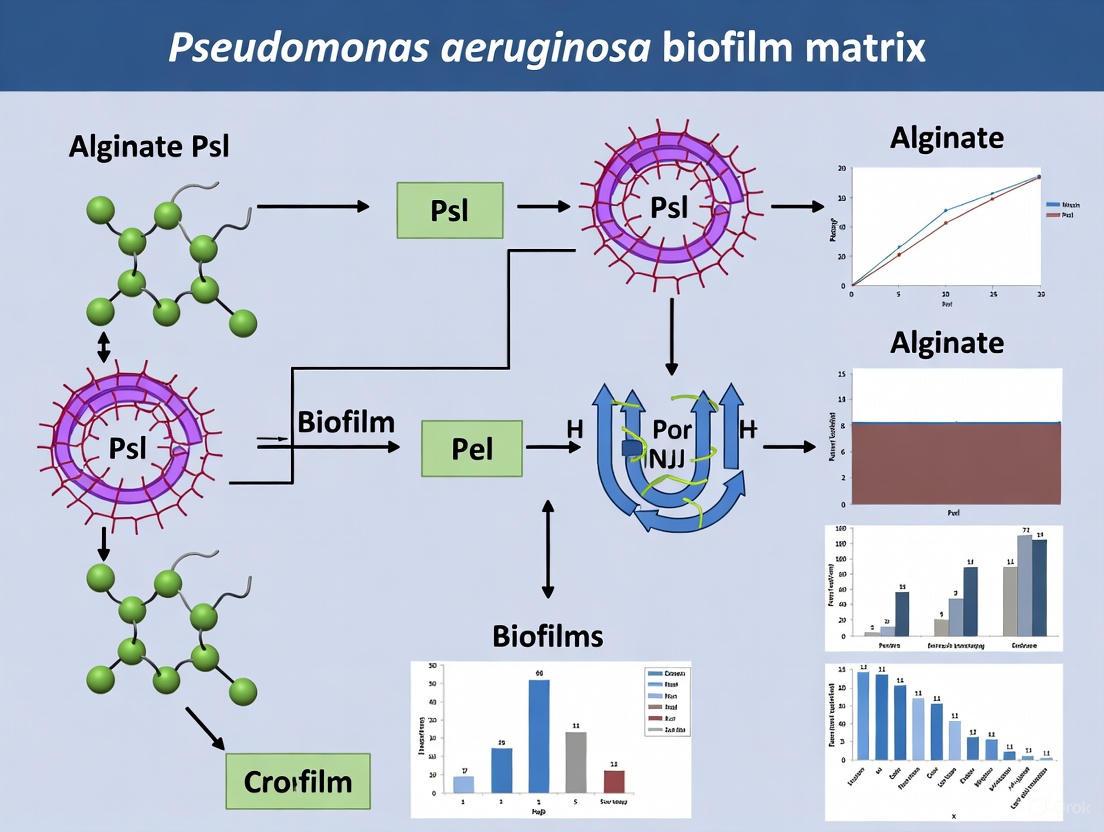
The chemical identities of alginate, Psl, and Pel—defined by their distinct monosaccharide compositions, linkages, and charge properties—directly determine their unique and often synergistic roles in constructing the P. aeruginosa biofilm matrix. Alginate forms a protective anionic hydrogel, Psl acts as a critical neutral scaffold for attachment and structure, and Pel provides cationic cross-linking functionality. This detailed understanding of the "matrixome" provides a critical foundation for future research. The experimental frameworks and reagents outlined here will empower ongoing investigations. Furthermore, the defined chemical structures and functional hierarchies of these exopolysaccharides illuminate promising targets for novel anti-biofilm strategies, such as enzymatic degradation and inhibition of biosynthesis, offering a pathway to overcome the formidable resistance posed by these bacterial communities.
This technical guide details the biosynthetic pathways of the three major exopolysaccharides—alginate, Psl, and Pel—produced by Pseudomonas aeruginosa. As critical components of the biofilm matrix, these polymers are central to the bacterium's resilience and pathogenicity. This whitepaper synthesizes current mechanistic models, comparing the gene operons, biosynthetic machinery, and export systems for each polymer. Framed within the broader context of biofilm matrix research, it provides structured data, experimental protocols, and visual tools to aid researchers and drug development professionals in targeting these pathways for therapeutic intervention.
Pseudomonas aeruginosa is an opportunistic pathogen whose adaptability stems partly from its capacity to produce extracellular polysaccharides, forming a protective biofilm matrix [10]. This matrix provides enhanced tolerance to desiccation, oxidizing agents, and host defenses [10]. Among these polymers, three secreted polysaccharides are predominant: alginate, Psl, and Pel [10] [4].
Most individual P. aeruginosa strains produce one secreted polysaccharide predominantly, though they possess the genetic capacity for all three [10]. The production of these exopolysaccharides is a key survival strategy, with significant implications in chronic infections, such as those in the lungs of cystic fibrosis patients, where the mucoid phenotype, characterized by alginate overproduction, is a marker of poor prognosis [10] [11]. Understanding their distinct biosynthetic routes—from gene regulation to polymer assembly and export—is crucial for developing anti-biofilm strategies.
Alginate Biosynthesis and Export
Alginate is the best-characterized exopolysaccharide in P. aeruginosa, particularly due to its role in chronic cystic fibrosis infections [10]. It is a high molecular weight, acidic polymer composed of non-repeating subunits of β-1,4-linked D-mannuronic acid and its C-5 epimer L-guluronic acid, with mannuronic acid residues being selectively O-acetylated [10] [11].
The Alginate Gene Operon and Multiprotein Complex
Proteins required for alginate biosynthesis are encoded by the algD operon (algD, alg8, alg44, algK, algE, algG, algX, algL, algI, algJ, algF, algA), with algC located elsewhere on the chromosome [11]. Experimental evidence confirms that these proteins form a trans-envelope multiprotein complex spanning the inner to outer membrane, ensuring coupled polymerization and translocation [11].
Table 1: Core Proteins in the Alginate Biosynthesis Complex
| Protein | Location | Key Function |
|---|---|---|
| AlgA, AlgD, AlgC | Cytoplasm | Conversion of fructose-6-P to GDP-mannuronic acid [11] |
| Alg8 | Inner Membrane | Glycosyltransferase; polymerizes GDP-mannuronic acid into polymannuronate chain [10] [11] |
| Alg44 | Inner Membrane | PilZ domain binds c-di-GMP; essential for polymerization [10] [11] |
| AlgG | Periplasm | C5-epimerase; converts D-mannuronic acid to L-guluronic acid [11] |
| AlgX | Periplasm | Interacts with AlgK and MucD; essential for secretion [11] |
| AlgK | Periplasm (OM-associated) | Tetratricopeptide repeats for protein-protein interactions; periplasmic scaffold [11] |
| AlgI, AlgJ, AlgF | Periplasm | Selective O-acetylation of mannuronic acid residues [11] |
| AlgL | Periplasm | Bifunctional; alginate lyase and periplasmic scaffold integrity [11] |
| AlgE | Outer Membrane | Forms an electropositive β-barrel secretion pore [11] |
Experimental Analysis of the Multiprotein Complex
Key experimental evidence for the alginate biosynthetic complex was demonstrated through mutual stability analysis, pulldown assays, and coimmunoprecipitation [11]. For instance, deleting algK in an alginate-overproducing strain (PDO300) destabilized Alg8 and Alg44 and interfered with alginate polymerization [11].
Protocol: Coimmunoprecipitation to Validate AlgE Interaction with AlgK
- Genetic Construct: Generate a FLAG-tagged variant of AlgE in a suitable plasmid vector [11].
- Strain Generation: Introduce the plasmid into a P. aeruginosa strain (e.g., PAO1ΔalgE) via conjugation or electroporation [11].
- Membrane Preparation: Grow the bacterial culture, harvest cells, and disrupt them using a French press or sonication. Isolate the membrane fraction via ultracentrifugation.
- Solubilization: Solubilize membrane proteins using a mild detergent (e.g., n-dodecyl-β-D-maltoside).
- Immunoprecipitation: Incubate the solubilized protein extract with anti-FLAG M2 affinity gel. Wash the resin extensively with a suitable buffer to remove non-specifically bound proteins.
- Elution and Analysis: Elute the bound proteins using FLAG peptide or SDS-PAGE loading buffer. Analyze the eluate by SDS-PAGE and Western blotting, probing for AlgK to confirm the interaction [11].
Diagram 1: The alginate biosynthesis and secretion multiprotein complex.
Psl Biosynthesis and Assembly
The Psl polysaccharide is a neutral polymer composed of a repeating pentamer containing D-mannose, L-rhamnose, and D-glucose [10]. It plays a critical role in cell-cell and cell-surface interactions during biofilm formation and forms a helical structure around the cell surface in strain PAO1 [10].
Predictive Model for Psl Biosynthesis
Psl biosynthesis is predicted to resemble the group 1 capsular polysaccharide pathway of Escherichia coli (e.g., the K30 antigen) [10] [4]. This model involves the assembly of pentameric subunits on an isoprenoid lipid carrier prior to transport and polymerization.
Table 2: Proposed Key Components in Psl Biosynthesis
| Component Type | Proposed Role in Psl Biosynthesis |
|---|---|
| Isoprenoid Lipid Carrier | Anchors the growing Psl pentameric subunit during assembly in the cytoplasm and for flip across the inner membrane [10]. |
| Glycosyltransferases | Enzymes encoded by the psl operon that sequentially add specific monosaccharides (D-mannose, L-rhamnose, D-glucose) to the lipid carrier [10]. |
| Wzx-like Translocase | Putative "flippase" that transports the lipid-linked Psl subunit from the cytoplasmic to the periplasmic face of the inner membrane [10]. |
| Wzy-like Polymerase | Putative polymerase that links the individual lipid-bound Psl subunits into a longer polymer in the periplasm [10]. |
| OPX (Outer Membrane Polysaccharide Export) Protein | Putative protein for the translocation of the polymer across the outer membrane, potentially forming a channel [10]. |
The psl operon (PA2231–PA2242 in PAO1) encodes the proteins for this pathway. Its identification via reverse genetics revealed its critical role in biofilm formation [10] [4].
Pel Biosynthesis and Modification
The Pel polysaccharide is a positively charged, glucose-rich polymer that is distinct from cellulose and crucial for pellicle formation and biofilm structure in strains like PA14 [10] [12]. Recent structural studies have significantly advanced the understanding of its biosynthesis.
The Pel Operon and a Unique Modification-Export System
The pel operon consists of seven genes [10]. A key feature of Pel biosynthesis is the coordinated action of the periplasmic proteins PelA and PelB for polymer modification and export [12].
- PelA is a bifunctional enzyme with four distinct domains. It possesses:
- A hydrolase domain with endo-α-1,4-N-acetylgalactosaminidase activity.
- A deacetylase domain, identified as the founding member of a new carbohydrate esterase family (CE21), which partially deacetylates the α-1,4 linked N-acetylgalactosamine (GalNAc) polymer [12].
- PelB is an integral outer membrane protein that likely forms a porin for polymer export [12].
The interaction between PelA and PelB is critical. AlphaFold2 modeling of the PelAB complex suggests a model where PelB guides the Pel polysaccharide to the deacetylase domain of PelA and subsequently to its own porin domain for export [12]. This interaction enhances the deacetylation efficiency of GalNAc oligosaccharides, and its perturbation can lead to less efficient deacetylation and increased Pel hydrolysis [12].
Diagram 2: Proposed Pel modification and export via the PelA-PelB complex.
Comparative Analysis of Biosynthetic Pathways
The three exopolysaccharides utilize distinct biosynthetic strategies, which can be broadly classified into two mechanistic classes.
Table 3: Comparative Biosynthetic Mechanisms of P. aeruginosa Exopolysaccharides
| Feature | Alginate | Psl | Pel |
|---|---|---|---|
| Chemical Composition | Acidic; D-mannuronic acid, L-guluronic acid [10] | Neutral; D-mannose, L-rhamnose, D-glucose pentamer [10] | Positively charged; partially deacetylated α-1,4-linked N-acetylgalactosamine [12] |
| Lipid Carrier Dependence | Independent (similar to cellulose) [10] | Dependent (isoprenoid lipid carrier) [10] | Predicted to be independent [10] |
| Polymerization Site | Inner membrane face by Alg8/Alg44 [10] [11] | Periplasm (Wzy-dependent) [10] | Not fully characterized, potentially inner membrane [10] |
| Key Regulatory Molecule | c-di-GMP (binds to Alg44) [11] | Not specified in results | Not specified in results |
| Periplasmic Modification | O-acetylation (AlgIJF), Epimerization (AlgG) [11] | Not specified in results | Partial deacetylation (PelA/CE21) [12] |
| Export Complex | Trans-envelope multiprotein complex (AlgGKXLE, etc.) [11] | OPX-like export system [10] | PelB porin guided by PelA [12] |
The Scientist's Toolkit: Key Research Reagents and Methodologies
Table 4: Essential Research Reagents and Materials for Pathway Analysis
| Reagent / Material | Function / Application in Research |
|---|---|
| Isogenic Knockout Mutants (e.g., PDO300ΔalgK) | To investigate the role of specific genes in polysaccharide biosynthesis, polymerization, and complex stability via mutual stability analysis [11]. |
| Complementation Plasmids (e.g., pBBR1MCS-5::algK) | For trans-complementation of knockout mutants to confirm gene function and rule out polar effects [11]. |
| Epitope-Tagged Protein Variants (e.g., AlgK-6xHis, FLAG-AlgE) | For protein-protein interaction studies using pulldown assays (His-tag) and coimmunoprecipitation (FLAG-tag) [11]. |
| N-Acetyl Cysteine (NAC) | A matrix-penetrating antimicrobial used to kill biofilm cells while leaving the matrix intact, allowing study of the remnant matrix's physical properties and role in recolonisation [13]. |
| Particle Tracking Microrheology (PTM) | A non-invasive technique using embedded microparticles to measure the local viscoelastic properties and mechanical stability of biofilms without structural disruption [13]. |
| AlphaFold2 Modeling | A computational tool used to predict the 3D structure of proteins and protein complexes, such as the interaction between PelA and PelB, providing mechanistic insights [12]. |
The biofilm matrix of Pseudomonas aeruginosa is a critical determinant of its success as an opportunistic pathogen, contributing significantly to chronic infections in settings such as cystic fibrosis (CF) lungs, chronic wounds, and on medical implants [14] [15]. This extracellular matrix provides structural integrity and confers formidable resistance to both antibiotics and host immune defenses [3] [16]. While the matrix is a composite of extracellular DNA, proteins, and lipids, the exopolysaccharides (EPS) serve as its foundational scaffold. P. aeruginosa is genetically equipped to produce three principal exopolysaccharides: alginate, Psl, and Pel [17] [18]. Each polysaccharide possesses a unique chemical structure and fulfills distinct, and sometimes overlapping, physiological roles—from initial surface attachment to the maintenance of complex three-dimensional community architecture and immune evasion [14] [19] [3]. This whitepaper synthesizes current research to delineate the primary roles of these EPS components, framing them within the broader context of targeted therapeutic interventions aimed at dismantling the protective biofilm barrier.
Structural Characteristics and Biosynthesis
The distinct chemical nature of each exopolysaccharide dictates its specific function within the biofilm matrix.
- Psl: Psl is a neutral, branched pentasaccharide composed of repeating units of D-mannose, D-glucose, and L-rhamnose in a 3:1:1 ratio [19] [16]. Its biosynthesis is directed by the 15-gene pslA-O operon. PslA is an inner membrane glycosyltransferase, while PslB is involved in producing the sugar-nucleotide precursor GDP-mannose [14]. Psl exists in both a cell-associated form, which forms a helical array on the bacterial surface, and a soluble form released into the environment [19].
- Pel: Pel is a cationic linear polymer composed of partially de-N-acetylated α-1,4-linked N-acetylgalactosamine and N-acetylglucosamine residues [14] [9] [16]. The positive charge, acquired through partial deacetylation in the periplasm, is a key feature that allows it to interact ionically with negatively charged molecules like extracellular DNA (eDNA) [9] [16]. The pelA-G operon encodes its synthesis and export machinery, with PelF serving as a cytoplasmic glycosyltransferase [14].
- Alginate: Alginate is a linear, anionic copolymer of β-D-mannuronic acid and its C5 epimer, α-L-guluronic acid [14] [19]. It is heavily O-acetylated, which contributes to its properties. The alg gene cluster, particularly alg8, which encodes a glycosyltransferase, is essential for its production [14]. Alginate is the hallmark exopolysaccharide of mucoid P. aeruginosa variants frequently isolated from the airways of CF patients [19].
Table 1: Core Characteristics of P. aeruginosa Exopolysaccharides
| Polysaccharide | Chemical Nature | Monosaccharide Composition | Key Biosynthetic Genes/Proteins |
|---|---|---|---|
| Psl | Neutral, branched | D-mannose, D-glucose, L-rhamnose [19] | pslA-O operon; PslA (glycosyltransferase), PslB [14] |
| Pel | Cationic, linear | Partially deacetylated N-acetylgalactosamine and N-acetylglucosamine [14] [16] | pelA-G operon; PelF (glycosyltransferase) [14] |
| Alginate | Anionic, linear | β-D-mannuronic acid, α-L-guluronic acid [14] | alg gene cluster; Alg8 (glycosyltransferase) [14] |
Functional Roles in Biofilm Development and Pathogenesis
Psl: The Master Mediator of Attachment and Initial Structure
Psl plays a dominant and multi-faceted role in the early and middle stages of biofilm formation. It is critically involved in surface attachment, serving as a primary adhesin that allows cells to initiate contact with substrates [17] [18]. Beyond attachment, Psl is a key architectural component. It forms a fibrous matrix that connects cells, guiding the formation of microcolonies and contributing significantly to the biomass, thickness, and elasticity of the mature biofilm [14]. Phenotypic studies demonstrate that deletion of the pslAB genes results in a severe defect, with biofilm levels dropping to less than one-third of the wild-type and the biofilm becoming significantly more rigid and less elastic [14]. Furthermore, Psl facilitates immune evasion by preventing complement deposition and opsonization in human serum, thereby protecting bacteria from phagocytic killing [19]. It also stimulates a proinflammatory response in the lung, which, while causing damage, can paradoxically aid in reducing bacterial colonization, indicating a complex host-pathogen interaction [19].
Pel: A Redundant yet Crucial Structural and Protective Scaffold
The role of Pel is highly strain-dependent but becomes critical when Psl is absent. In strains like PA14, Pel serves as the primary structural polysaccharide, essential for maintaining cell-to-cell interactions and the three-dimensional architecture of biofilms [17] [3]. It confers a redundant function in many strains; for instance, in a Psl-deficient PAO1 background, the biofilm is initially poor, but over time, mutations can arise that upregulate Pel expression, allowing biofilm formation to be restored [17]. This functional redundancy highlights the evolutionary advantage of possessing multiple polysaccharide systems. A second key role of Pel is in antibiotic tolerance. Pel specifically enhances resistance to aminoglycoside antibiotics (e.g., tobramycin) within biofilm populations, likely through cationic sequestration of these positively charged drug molecules [3] [16]. Its positive charge also enables it to cross-link with eDNA, forming a stable composite matrix that protects the eDNA from degradation by nucleases like DNase I [16].
Alginate: A Shield for Immune Evasion in Chronic Infections
Alginate's primary role is not in initial biofilm formation in non-mucoid strains but in protecting established biofilms from the host immune system. Its production is the defining characteristic of the mucoid phenotype, which is frequently selected for in the chronic, inflamed CF lung environment [19]. Alginate acts as a physical barrier, inhibiting phagocytosis by immune cells such as neutrophils and macrophages [19] [15]. It also provides protection against damage from reactive oxygen species (ROS) [19]. By forming a hydrated, gelatinous layer around bacterial aggregates, alginate sterically hinders the access of immune effectors and some antimicrobials, thereby promoting bacterial persistence [15] [16].
Table 2: Primary Functional Roles and Phenotypic Consequences of EPS Deletion
| Polysaccharide | Primary Demonstrated Roles | Key Phenotypes of Mutant Strains | Contextual Dependence & Redundancy |
|---|---|---|---|
| Psl | Surface attachment, biofilm structural scaffolding, biofilm elasticity, immune evasion (anti-opsonization) [14] [19] [17] | Severe reduction in attachment and biofilm biomass (to <1/3 of WT); increased biofilm rigidity [14] [17] [18] | Primary structural polysaccharide in many strains (e.g., PAO1); critical for most isolates' surface attachment [17] |
| Pel | Structural scaffolding (cell-cell interactions), antibiotic tolerance (aminoglycosides), functional redundancy [17] [3] [16] | Reduced biofilm biomass and loss of pellicle in Pel-dependent strains (e.g., PA14); increased aminoglycoside susceptibility [17] [3] | Can serve as primary scaffold (e.g., PA14); often redundant, can compensate for Psl loss upon evolutionary adaptation [17] |
| Alginate | Immune evasion (inhibition of phagocytosis, protection from ROS) [19] [15] | Negligible impact on biofilm formation in non-mucoid lab strains (e.g., PAO1) [14] | Dominant EPS in mucoid variants, particularly in chronic CF lung infections [19] |
Experimental Approaches for Functional Analysis
A combination of phenotypic, molecular, and omics techniques is employed to dissect the contributions of each EPS.
Phenotypic Characterization of Biofilms
- Crystal Violet Staining for Biomass Quantification: This classic assay measures total biofilm formation on abiotic surfaces (e.g., plastic or glass) after static incubation. It is used to compare biofilm-forming capacity between wild-type and isogenic EPS mutant strains [14] [3]. For example, a ΔpslA mutant typically shows a dramatic reduction (e.g., ~85%) in crystal violet staining compared to the wild-type [18].
- Microscopic Imaging for Architectural Analysis:
- Scanning Electron Microscopy (SEM): Provides high-resolution, topological images of the biofilm surface, revealing differences in density and structure. The ΔpslAB mutant biofilm appears sparse and underdeveloped compared to the dense wild-type structure [14].
- Confocal Laser Scanning Microscopy (CLSM): Allows for non-invasive, three-dimensional visualization of live biofilms, often in flow-cell reactors. It is used to quantify key architectural parameters such as biovolume, average thickness, and roughness using software like COMSTAT [14] [17]. A ΔpslAB mutant, for instance, exhibits approximately half the biomass and thickness of the wild-type, with a significant increase in roughness [14].
- Quartz Crystal Microbalance with Dissipation (QCM-D) Monitoring: This label-free technique monitors biofilm formation in real-time by measuring changes in the frequency (Δf, related to mass) and dissipation (ΔD, related to viscoelasticity) of an oscillating sensor crystal [14]. Biofilms with Psl display a characteristic decrease in frequency and increase in dissipation, indicating the formation of a thick, viscoelastic layer. In contrast, a ΔpslAB mutant shows minimal changes, indicating poor growth and the formation of a more rigid biofilm [14].
Molecular and Multi-Omics Techniques
- Transcriptomics and Metabolomics: Integrated multi-omics approaches reveal the internal regulatory mechanisms governing biofilm formation. For example, transcriptomic analysis of a ΔpslAB mutant reveals widespread dysregulation, including disruptions in chemotaxis, quorum sensing, and c-di-GMP/amino acid metabolism, underscoring Psl's central role as a regulatory hub [14].
- Mutant Strain Construction: In-frame deletion mutants of key biosynthetic genes (e.g., pslD for Psl, pelF for Pel, and alg8 for alginate) are fundamental tools for establishing genotype-phenotype relationships [14] [17] [18]. The use of non-polar mutations is critical to avoid polar effects on downstream genes.
Diagram 1: Functional integration of Psl, Pel, and Alginate in biofilm development and pathogenesis. Psl (yellow) is involved in all stages, from initial attachment to immune evasion. Pel (green) contributes to structural integrity and specific antibiotic tolerance. Alginate (red) is primarily deployed for immune evasion, facilitating chronic persistence. A key regulatory feedback loop involves c-di-GMP, which is elevated in mature biofilms and further stimulates Psl production.
The Scientist's Toolkit: Key Reagents and Experimental Models
Table 3: Essential Research Reagents and Models for EPS Studies
| Tool/Reagent | Function/Description | Experimental Application |
|---|---|---|
| Anti-Psl Monoclonal Antibodies | Antibodies specifically binding to Psl epitopes [19] | Visualization and localization of Psl within biofilms via immunofluorescence/CLSM [19] |
| PslG Hydrolase | Glycoside hydrolase that enzymatically degrades the Psl polysaccharide [19] [20] | Targeted disruption of Psl-dependent biofilms; testing Psl's essential role in biofilm structure and antibiotic tolerance [19] |
| Alginate Lyase | Enzyme that cleaves alginate polymers [20] | Degradation of alginate in mucoid biofilms; used to study alginate's role in immune evasion and to potentiate antibiotic efficacy [20] |
| Isogenic Mutant Strains (e.g., ΔpslA, ΔpelF, Δalg8) | Strains with precise genetic deletions in specific EPS biosynthetic genes [14] [17] [18] | Fundamental for comparative phenotyping (vs. wild-type) to define the specific function of each EPS [14] [18] |
| Flow Cell Biofilm Reactors | Devices allowing continuous nutrient flow for biofilm growth under shear stress [17] [3] | Cultivation of biofilms for real-time, non-destructive imaging and analysis using CLSM [17] |
| QSense QCM-D Analyzer | Instrument for real-time, label-free monitoring of biofilm mass and viscoelastic properties [14] | Quantitative assessment of biofilm development kinetics and mechanical properties in response to genetic or chemical perturbations [14] |
Discussion and Future Perspectives
The delineation of non-redundant primary roles for Psl (attachment), Pel (structural scaffolding with functional redundancy), and alginate (immune evasion) provides a clear conceptual framework for both understanding biofilm pathobiology and designing targeted anti-biofilm strategies. The hierarchical importance of Psl, as revealed by multi-omics studies showing its deletion causes widespread metabolic and regulatory disruption, positions it as a high-priority therapeutic target [14]. Furthermore, the strain-specificity and functional redundancy between Pel and Psl underscore the adaptability of P. aeruginosa and suggest that combination therapies targeting multiple EPS components may be necessary for effective biofilm eradication [17] [16].
Emerging therapeutic approaches directly stem from this functional understanding. The use of glycoside hydrolases like PslG to dismantle the Psl matrix has shown promise in pre-clinical models, significantly reducing biofilm biomass and potentiating the activity of conventional antibiotics [19] [20]. Similarly, alginate lyase is being investigated to break down the protective shield of mucoid strains [20]. As research progresses, the integration of these EPS-targeting agents with standard-of-care antibiotics represents a promising frontier for overcoming the recalcitrance of P. aeruginosa biofilm infections, with the ultimate goal of improving clinical outcomes for affected patients.
Pseudomonas aeruginosa thrives in diverse environments and causes devastating chronic infections in immunocompromised individuals and cystic fibrosis (CF) patients by forming biofilms—structured communities of bacteria encased in an extracellular matrix [17]. The biofilm matrix provides structural integrity and protects embedded bacteria from antibiotics and host immune responses [16]. This protective matrix contains three key exopolysaccharides (EPS)—alginate, Psl, and Pel—each with distinct chemical properties and functional roles [17] [7]. While non-mucoid strains typically utilize Pel or Psl as primary structural components, mucoid variants often overproduce alginate, a hallmark of chronic CF lung infections [17] [16].
Despite the significant biosynthetic cost, P. aeruginosa maintains the genetic capacity to produce multiple EPS types, suggesting an evolutionary advantage to this redundancy. This review examines how strain-specific EPS expression patterns and functional redundancy between Pel and Psl provide P. aeruginosa with remarkable adaptability in biofilm formation across diverse environments and under selective pressures. Understanding these mechanisms is crucial for developing effective anti-biofilm strategies against this persistent pathogen.
Structural and Functional Diversity of P. aeruginosa Exopolysaccharides
The three major exopolysaccharides produced by P. aeruginosa differ significantly in their chemical composition, biosynthetic pathways, and specific roles during biofilm development.
Table 1: Characteristics of Major P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Genetic Locus | Primary Functions in Biofilms | Charge |
|---|---|---|---|---|
| Psl | Neutral pentasaccharide (D-mannose, D-glucose, L-rhamnose) [17] | psl (15 genes, pslA-O) [16] | Surface attachment, structural scaffold, cell-cell connections, antibiotic sequestration [17] [16] | Neutral |
| Pel | Cationic polymer of partially de-N-acetylated galactosamine and N-acetyl galactosamine [16] | pel (7 genes, pelA-G) [17] | Pellicle formation, cell-cell interactions, structural scaffold, eDNA binding [17] [7] | Positive |
| Alginate | Linear polyanionic polymer of uronic acids [7] | alg (alginate operon) | Biofilm architecture, cell viability protection, decreased antibiotic susceptibility [7] | Negative |
Psl: A Multifunctional Adhesin and Scaffold
Psl plays crucial roles throughout biofilm development. This neutral polysaccharide forms helical arrangements on the bacterial surface that facilitate initial attachment to both biotic and abiotic surfaces [16]. During surface exploration, P. aeruginosa deposits Psl trails that act as chemical signals guiding subsequent cell movement and microcolony formation [21]. In mature biofilms, Psl provides structural integrity by cross-linking with the adhesin CdrA and extracellular DNA (eDNA) [16]. It also contributes to antibiotic tolerance by physically limiting antibiotic penetration and sequestering certain antimicrobial agents [16].
Pel: The Cationic Scaffold for Cell-Cell Interactions
Pel functions as a key structural component in pellicles (biofilms at the air-liquid interface) and in cell-cell adhesion during biofilm formation [17] [7]. Its positive charge enables electrostatic interactions with negatively charged eDNA, forming ionic cross-links that strengthen the biofilm matrix [16]. These Pel-eDNA complexes protect the genetic material from degradation by DNases, potentially limiting the efficacy of nuclease-based therapies [16]. Pel also contributes to antibiotic tolerance, particularly against cationic aminoglycosides like tobramycin, through sequestration mechanisms [16].
Alginate: The Protective Shield in Chronic Infections
Alginate production is typically associated with mucoid variants of P. aeruginosa that emerge during chronic CF lung infections [17]. This polysaccharide forms a gelatinous coating that promotes bacterial aggregation and protects embedded cells from antibiotics and host immune effectors [7] [16]. Alginate-overproducing strains develop biofilms with distinct architectures characterized by finger-like microcolonies and enhanced structural stability [7].
Strain-Specificity in Exopolysaccharide Utilization
Research has revealed remarkable strain-to-strain variability in EPS dependence among P. aeruginosa isolates, reflecting adaptation to specific environmental niches and selective pressures.
Classification of Strains by EPS Dependence
Table 2: Functional Classification of P. aeruginosa Strains Based on EPS Dependence
| Strain Class | Primary Structural EPS | Attachment Dependence | Representative Strains | Key Characteristics |
|---|---|---|---|---|
| Class I | Psl | Psl-dependent | PAO1 [17] | Psl critical for attachment and mature biofilm structure |
| Class II | Pel | Psl-dependent | PA14 [17] | Psl required for attachment, Pel for mature biofilm structure |
| Class III | Pel and Psl | Psl-dependent | Clinical and environmental isolates [17] | Functional redundancy between Pel and Psl in mature biofilms |
| Class IV | Alginate | Psl-dependent | Mucoid CF isolates [17] [7] | Alginate as dominant EPS, often with retained Psl function |
The variation in EPS dependence is exemplified by the differential behavior of commonly studied laboratory strains. PAO1 relies primarily on Psl for both initial attachment and mature biofilm structure, while PA14 (which has a natural deletion in the psl operon) depends on Pel for mature biofilm development, despite both strains requiring Psl for initial surface attachment [17]. This fundamental difference highlights how even closely related strains can employ distinct EPS strategies for biofilm formation.
Molecular Basis for Strain Variation
The strain-specific EPS profiles arise from complex regulatory networks that control pel and psl expression. Key regulators include:
- FleQ: Acts as a transcriptional repressor for both pel and psl operons, with repression relieved by the secondary messenger c-di-GMP [17].
- c-di-GMP: Serves as a positive regulator of Pel synthesis through direct binding to PelD and modulates transcription of both pel and psl operons [17].
- RsmA: An RNA-binding protein that inhibits psl translation, providing post-transcriptional control [17].
- Quorum Sensing: Influences pel and psl expression through cell-density dependent signaling pathways [17].
These regulatory mechanisms allow P. aeruginosa to fine-tune EPS production in response to environmental cues, contributing to the observed strain-to-strain variability.
Figure 1: Regulatory Network Controlling Pel and Psl Expression. Multiple environmental and genetic factors converge to regulate the production of key exopolysaccharides in P. aeruginosa biofilms.
Functional Redundancy Between Pel and Psl
The capacity to produce both Pel and Psl provides P. aeruginosa with a backup system that preserves biofilm formation capability when one EPS system is compromised.
Experimental Evidence for Redundancy
Key experiments demonstrating functional redundancy include:
Mutational Analysis in Clinical Isolates: Studies comparing clinical and environmental isolates revealed that while Psl is critical for initial surface attachment across most strains, the contribution of Pel and Psl to mature biofilm structure shows significant variability [17]. Some strains could maintain biofilm integrity through Pel production even when Psl was knocked out, and vice versa.
Evolution Experiments: When a PAO1 psl mutant strain (primarily dependent on Psl for biofilm formation) was cultivated for extended periods, it acquired mutations that upregulated Pel polysaccharide production, compensating for the lack of Psl and restoring biofilm formation capacity [17]. This demonstrates the evolutionary advantage of maintaining both systems.
Genetic Compensation: Studies have shown that the absence of one EPS can enhance production of another. For instance, a lack of Psl enhances Pel production, while absence of Pel can increase alginate production in certain genetic backgrounds [7]. This compensatory regulation ensures biofilm integrity is maintained.
Molecular Mechanisms of Redundancy
The molecular basis for redundancy involves shared regulatory pathways and structural complementarity:
- Shared Regulatory Elements: Both pel and psl operons respond to similar regulatory signals, particularly c-di-GMP, allowing coordinated expression in response to environmental conditions [17].
- Structural Complementarity: While Psl and Pel have distinct chemical properties, both can provide structural scaffolding in mature biofilms [17]. The neutral Psl and cationic Pel can each interact with other matrix components to maintain biofilm architecture.
- Cross-talk with Other Matrix Components: Both Pel and Psl interact with eDNA and matrix proteins to form the biofilm scaffold, though through different mechanisms—Pel via electrostatic interactions and Psl through specific binding partners [7] [16].
Figure 2: Functional Redundancy Mechanism Between EPS Systems. P. aeruginosa maintains biofilm functionality through compensatory regulation when primary EPS systems are compromised.
Experimental Methodologies for EPS Research
Genetic Manipulation of EPS Pathways
Construction of Mutant Strains: Researchers generate in-frame deletions of key biosynthetic genes (pelF for Pel, pslD for Psl, alg8 for alginate) using allelic replacement strategies [17] [7]. For example, to create a pelF mutant, two regions of the pelF gene are amplified by PCR, hydrolyzed with restriction enzymes, ligated together, and inserted into a vector such as pGEM-T Easy [7]. A gentamicin resistance cassette is inserted, and the construct is transferred into P. aeruginosa via electroporation. Transformants are selected using antibiotic resistance, and gene replacement is confirmed via PCR [7].
Complemented Strains: For functional validation, mutant strains are complemented by introducing the wild-type gene on a broad-host-range vector such as pBBR1-MCS5 [7]. The gene of interest is amplified from chromosomal DNA, inserted into the vector, and transferred into the mutant strain.
Biofilm Assessment Methods
Microtiter Dish Assay: This static method measures biofilm formation at the air-liquid interface after incubation. Bacterial attachment is quantified using crystal violet staining or directly measuring biofilm biomass [17].
Flow-Cell Reactor with CLSM: Biofilms grown under continuous flow conditions in specialized chambers are visualized using confocal laser scanning microscopy (CLSM) [17]. This allows real-time observation of biofilm architecture, microcolony development, and spatial organization of different matrix components.
Bulk Rheological Measurements: The mechanical properties of biofilms are quantified using stress-controlled rheometers with parallel-plate geometry [22]. Biofilms are gently scraped onto the rheometer plate, trimmed to fill a 500μm gap, and subjected to oscillatory shear stress to measure storage modulus (G'), loss modulus (G''), and yield stress [22].
EPS-Specific Enzymatic Tools
Targeted Matrix Disruption: Researchers employ specific enzymes to dissect EPS functions: alginate lyase degrades alginate via β-elimination at β-1,4 bonds; Pel-specific glycosidases target Pel; and Psl-specific glycosidases degrade Psl [22]. DNase I cleaves eDNA, which interacts particularly with Pel [22].
Treatment Protocols: Enzymes are applied by spreading 50-100μL of treatment solution over biofilm lawns grown on agar plates, incubating upright for one hour at 37°C before analysis [22].
Table 3: Essential Research Reagents for EPS Studies
| Reagent Category | Specific Examples | Research Applications | Key Functions |
|---|---|---|---|
| EPS-Deficient Mutants | PAO1 ΔpslD, PAO1 ΔpelF, PA14 Δpel [17] | Functional analysis of specific EPS | Genetic dissection of EPS-specific roles |
| EPS-Overproducing Strains | PAO1 ΔwspF (hyperbiofilm former) [22] [23] | Studies of EPS production regulation | Model for Rugose Small Colony Variants |
| EPS-Specific Enzymes | Alginate lyase, Psl glycosidase, Pel glycosidase [22] | Targeted matrix disruption | Functional dissection of EPS components |
| c-di-GMP Modulators | Chemical inducers/inhibitors of diguanylate cyclases/phosphodiesterases [17] | Regulation studies of EPS expression | Manipulation of key signaling pathway |
| Fluorescent Reporters | Psl-gfp, Pel-gfp transcriptional fusions [17] | Spatial localization of EPS production | Visualization of EPS distribution in biofilms |
Therapeutic Implications and Future Directions
Exploiting EPS Specificity for Targeted Therapies
Understanding strain-specific EPS dependencies opens avenues for precision anti-biofilm approaches:
Enzyme-Based Treatments: The efficacy of enzymatic biofilm disruption depends on matching the enzyme to the dominant EPS produced by the infecting strain [22]. For instance, alginate lyase most effectively compromises the mechanics of alginate-dominant biofilms, while DNase I has greater impact on Pel-dominant biofilms where Pel-eDNA interactions are critical [22].
Phage Therapy Innovations: Some bacteriophages have evolved to exploit biofilm matrix components, such as Psl-dependent phages that bind to and degrade this polysaccharide to access bacterial cells [23]. Directed evolution of phages in biofilms has yielded mutants with enhanced efficacy through improved recognition of bacterial surface structures [24].
Antibiotic Adjuvants: EPS-targeting agents can potentiate conventional antibiotics. For example, Psl-degrading enzymes increase susceptibility to tobramycin in Psl-dependent biofilms [16].
Challenges in Clinical Translation
Several factors complicate therapeutic strategies targeting EPS:
Environmental Influence on Matrix Composition: Biofilms grown in different environments (in vitro vs. in vivo) show substantial differences in matrix composition, affecting treatment efficacy [22]. Enzymes highly effective against in vitro biofilms may show reduced activity in animal models [22].
Adaptive Compensation: Therapeutic targeting of one EPS may select for strains that upregulate alternative EPS, maintaining biofilm integrity through redundant systems [17].
Strain Diversity: The significant strain-to-strain variability in EPS dependence necessitates personalized approaches based on the EPS profile of the infecting strain [17].
P. aeruginosa invests in multiple exopolysaccharides as an evolutionary strategy to ensure biofilm formation across diverse environments and under varying selective pressures. The strain-specificity in EPS utilization reflects adaptation to particular niches, while the functional redundancy between Pel and Psl provides a safeguard against genetic or environmental disruption of either system. This sophisticated portfolio of matrix components, regulated by complex signaling networks, enhances the resilience of P. aeruginosa biofilms and complicates therapeutic interventions. Future anti-biofilm strategies must account for both the strain-specific dependencies and compensatory capabilities inherent in this redundant system, potentially requiring combination approaches that target multiple EPS simultaneously or exploit the specific vulnerabilities of different strain classes.
The formation of biofilms is a key virulence factor for the opportunistic pathogen Pseudomonas aeruginosa, conferring high levels of resistance to antimicrobial agents and the host immune system. The extracellular polymeric substance (EPS) matrix, a critical biofilm component, is primarily composed of exopolysaccharides—alginate, Psl, and Pel. The production of these exopolysaccharides is intricately regulated by a complex network of intracellular and intercellular signaling systems. This review provides an in-depth analysis of how the secondary messenger bis-(3'-5')-cyclic dimeric guanosine monophosphate (c-di-GMP), quorum sensing (QS), and other signals orchestrate the biosynthesis of the EPS matrix. Framed within the context of alginate, Psl, and Pel research, this technical guide synthesizes current knowledge on regulatory mechanisms, presents quantitative data and experimental methodologies, and visualizes signaling pathways to serve researchers and drug development professionals in designing novel anti-biofilm strategies.
Pseudomonas aeruginosa is a model organism for biofilm research and a formidable opportunistic pathogen responsible for chronic infections in immunocompromised individuals and cystic fibrosis (CF) patients [25] [26]. Its resilience is largely attributed to the biofilm lifestyle, where bacterial communities are encased in a self-produced EPS matrix. This matrix constitutes over 90% of the biofilm biomass and acts as a structural scaffold and a protective barrier [25]. The most critical structural components of the P. aeruginosa EPS are three exopolysaccharides: alginate, Psl, and Pel.
Alginate is a high molecular weight, acetylated polymer composed of non-repeating subunits of D-mannuronic acid and its C-5 epimer L-guluronic acid [27] [4]. It is predominantly produced by mucoid variants of P. aeruginosa that are frequently isolated from the lungs of CF patients, and its overproduction correlates with the transition from acute to chronic infection [25] [4]. Alginate contributes to biofilm maturation and protects bacteria from phagocytosis and antibiotic penetration [25].
Psl is a neutral pentasaccharide containing D-mannose, L-rhamnose, and D-glucose [25] [4]. It is critical for initial surface attachment, cell-cell interactions, and maintaining the structural integrity of biofilms in non-mucoid strains like PAO1 [25] [28]. Psl forms a fiber-like matrix that enmeshes bacterial cells and has been visualized in a helical pattern around the cell surface during initial attachment [28]. Furthermore, Psl functions as a signaling molecule that stimulates the production of the intracellular second messenger c-di-GMP, creating a positive feedback loop for biofilm formation [26].
Pel is a glucose-rich, cationic exopolysaccharide [25] [26]. Its production is crucial for pellicle formation at the air-liquid interface and provides structural integrity to biofilms, particularly in strain PA14 [4] [26]. Pel also enhances tolerance to aminoglycoside antibiotics [25]. The precise biochemical structure of Pel remains undefined, though it is known to be distinct from alginate and Psl [4] [26].
The synthesis of these exopolysaccharides is not constitutive but is tightly regulated by a network of environmental and cellular signals. The following sections delve into the core regulatory systems controlling EPS production, with a focus on the central role of c-di-GMP and its integration with other signaling pathways.
The Central Role of Cyclic-di-GMP in EPS Regulation
c-di-GMP Metabolism and Basic Signaling Principles
The ubiquitous bacterial second messenger c-di-GMP is a central regulator of the transition from a motile, planktonic lifestyle to a sessile, biofilm-forming state [29] [30]. The intracellular concentration of c-di-GMP is determined by the opposing activities of two classes of enzymes:
- Diguanylate cyclases (DGCs): Synthesize c-di-GMP from two molecules of GTP. These enzymes are characterized by the presence of a conserved GGDEF domain [29] [31].
- Phosphodiesterases (PDEs): Degrade c-di-GMP. They feature either an EAL domain, which hydrolyzes c-di-GMP into linear 5'-phosphoguanylyl-(3'-5')-guanosine (pGpG), or an HD-GYP domain, which further degrades it into two GMP molecules [29].
P. aeruginosa possesses one of the largest sets of proteins involved in c-di-GMP metabolism, with genomes encoding 18 GGDEF domain proteins, 5 EAL domain proteins, 16 hybrid GGDEF-EAL proteins, and 3 HD-GYP domain proteins [29]. This abundance suggests a complex regulatory network capable of integrating diverse environmental signals to fine-tune cellular c-di-GMP levels.
A fundamental paradigm in c-di-GMP signaling is that high intracellular levels promote biofilm formation and repress motility, while low levels have the opposite effect [29] [30]. For instance, P. aeruginosa biofilms contain 75–110 pmol of c-di-GMP per mg of total cell extract, whereas planktonic cells contain less than 30 pmol mg⁻¹ [29]. This elevated c-di-GMP concentration stimulates the production of biofilm matrix components, including the exopolysaccharides Psl, Pel, and alginate.
Direct Activation of Exopolysaccharide Biosynthesis by c-di-GMP
A key mechanism by which c-di-GMP promotes EPS production is through the allosteric activation of polysaccharide biosynthesis complexes. Unlike transcriptional regulation, this post-translational control allows for rapid phenotypic changes in response to environmental cues.
Activation of Pel Biosynthesis: The PelD protein of P. aeruginosa was identified as a novel c-di-GMP receptor that directly mediates the regulation of Pel polysaccharide biosynthesis [32]. PelD contains a specific c-di-GMP binding site, and mutations in conserved residues critical for binding abolish Pel production without affecting the transcription of the pel operon [32]. This finding established that c-di-GMP can directly bind to components of the EPS biosynthesis machinery to activate polymer production.
Activation of Alginate and Psl Biosynthesis: While the precise mechanisms for alginate and Psl are less defined, it is established that elevated c-di-GMP levels boost their production [30]. For Psl, a positive feedback circuit exists whereby Psl itself stimulates the DGCs SiaD and SadC to produce more c-di-GMP, which in turn further enhances Psl synthesis [26]. The table below summarizes the regulatory relationships between c-di-GMP and the key exopolysaccharides.
Table 1: Regulation of P. aeruginosa Exopolysaccharides by c-di-GMP
| Exopolysaccharide | Role in Biofilm | Effect of High c-di-GMP | Key Regulatory Proteins / Mechanisms |
|---|---|---|---|
| Pel | Structural scaffold in PA14; pellicle formation; antibiotic tolerance. | Increased production. | Direct allosteric activation via PelD receptor [32]. |
| Psl | Initial attachment & structural integrity in PAO1; cell-cell adhesion; signaling. | Increased production. | Positive feedback loop stimulating DGCs SiaD & SadC [26]. |
| Alginate | Biofilm maturation in mucoid variants; protection from host defenses. | Increased production. | Regulation of alginate biosynthetic operon expression [30]. |
The following diagram illustrates the core signaling logic of c-di-GMP in promoting biofilm formation through the activation of EPS production.
Figure 1: Core c-di-GMP Signaling Logic. Environmental signals activate DGCs and inhibit PDEs, leading to a high intracellular c-di-GMP pool. High c-di-GMP allosterically activates EPS biosynthesis complexes and represses motility, promoting the biofilm lifestyle.
Integration with Quorum Sensing and Other Regulatory Systems
While c-di-GMP is a central regulator, EPS production is not controlled in isolation. It is integrated with the Quorum Sensing (QS) system, which allows bacterial populations to coordinate gene expression based on cell density.
The QS system in P. aeruginosa involves the production, release, and detection of acyl-homoserine lactone (AHL) signaling molecules. The LasI/LasR and RhlI/RhlR systems form a hierarchical regulatory network that controls the expression of hundreds of genes, including virulence factors and public goods [25]. While the direct transcriptional control of the alg, psl, and pel operons by QS is complex and strain-dependent, QS influences the biofilm matrix profoundly by regulating the production of other matrix components, such as extracellular DNA (eDNA) and lectins, which interact with and stabilize the exopolysaccharide network [25] [33].
Furthermore, a critical integration point exists between QS and c-di-GMP signaling. The QS system can regulate the expression of genes encoding DGCs and PDEs, thereby modulating the global c-di-GMP pool [29]. Conversely, certain c-di-GMP effectors can influence the production or activity of QS signals. This cross-talk creates a sophisticated regulatory circuit that ensures biofilm development is initiated and matured at the appropriate time and place.
Other regulatory inputs, such as nutritional cues (e.g., availability of carbon sources) and stress signals (e.g., oxidative stress, antibiotic exposure), also feed into this network, primarily by affecting the activity of specific DGCs and PDEs that contain sensory domains [30].
Table 2: Interplay Between c-di-GMP, Quorum Sensing, and Other Signals in EPS Regulation
| Regulatory System | Key Components | Interaction with EPS Regulation |
|---|---|---|
| c-di-GMP Signaling | DGCs (GGDEF), PDEs (EAL/HD-GYP) | Directly activates Pel via PelD; elevates Psl and alginate production; high levels promote biofilm matrix production. |
| Quorum Sensing (QS) | LasI/LasR, RhlI/RhlR, AHLs | Coordinates population-level behavior; can regulate expression of DGCs/PDEs; controls other matrix components (eDNA, proteins). |
| Nutritional/Stress Signals | Available nutrients, antibiotics, host defenses | Modulate activity of sensory domains on DGCs/PDEs, fine-tuning c-di-GMP levels and EPS production in response to the environment. |
The following diagram synthesizes this integrated regulatory network controlling Psl, Pel, and alginate production.
Figure 2: Integrated Regulatory Network for EPS Production. Environmental and host-derived signals influence the c-di-GMP pool by modulating the activity of specific DGCs and PDEs. Quorum sensing interacts with c-di-GMP signaling via cross-talk and feedback. The resulting high c-di-GMP level directly and indirectly upregulates the production of Psl, Pel, and alginate, leading to robust biofilm formation.
Quantitative Data and Experimental Analysis
Key Quantitative Findings
Research into the regulation of EPS has yielded crucial quantitative data that underpins our understanding of the system.
Table 3: Key Quantitative Data in EPS and c-di-GMP Research
| Parameter Measured | Quantitative Finding | Experimental Context / Strain | Significance / Interpretation |
|---|---|---|---|
| Intracellular c-di-GMP Concentration | Biofilms: 75-110 pmol/mg cell extract. Planktonic cells: <30 pmol/mg [29]. | P. aeruginosa biofilm vs. planktonic cultures. | Confirms correlation between high c-di-GMP and the biofilm lifestyle. |
| Enzymatic Potential for c-di-GMP Turnover | Genome encodes 18 GGDEF, 5 EAL, 16 GGDEF-EAL hybrid, and 3 HD-GYP domain proteins [29]. | P. aeruginosa PAO1 genome analysis. | Indicates a vast, complex network for fine-tuning c-di-GMP levels. |
| c-di-GMP Binding Affinity (PelD) | Identification of conserved residues (RXXD) required for c-di-GMP binding [32]. | In vitro binding assays with purified PelD protein. | Establishes PelD as a direct receptor for c-di-GMP in Pel activation. |
| Psl-Mediated c-di-GMP Feedback | Psl stimulates DGCs SiaD & SadC to produce more c-di-GMP [26]. | In vivo studies with psl mutants and c-di-GMP reporters. | Demonstrates a positive feedback loop that stabilizes the biofilm state. |
Essential Experimental Protocols
A deep understanding of this field requires familiarity with the key methodologies used to uncover these regulatory mechanisms.
Protocol 1: Measuring Intracellular c-di-GMP Levels
- Objective: To quantify the concentration of c-di-GMP in bacterial cells under different growth conditions (e.g., biofilm vs. planktonic).
- Methodology:
- Cell Extraction: Grow P. aeruginosa cultures to the desired phase. Harvest cells by rapid centrifugation and immediately extract nucleotides using a quenching solution like cold methanol or acetonitrile.
- Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS): Separate the extracted nucleotides using reverse-phase liquid chromatography. Use multiple reaction monitoring (MRM) on a triple-quadrupole mass spectrometer to specifically detect and quantify c-di-GMP based on its mass transition (e.g., 690>540).
- Normalization: Normalize the measured c-di-GMP concentration to the total protein content or cell dry weight of the sample to allow for comparison [29].
- Key Reagents: Cold methanol/acetonitrile, LC-MS grade solvents, c-di-GMP standard for calibration.
Protocol 2: Identifying Direct c-di-GMP Receptors (e.g., PelD)
- Objective: To demonstrate a direct protein-c-di-GMP interaction and its functional consequence.
- Methodology:
- Protein Purification: Clone and express the candidate receptor protein (e.g., PelD) in E. coli and purify it using affinity chromatography (e.g., His-tag purification).
- In Vitro Binding Assays:
- Isothermal Titration Calorimetry (ITC): Titrate a c-di-GMP solution into the purified protein solution. The heat change measured reveals the binding affinity (Kd), stoichiometry (N), and thermodynamic profile.
- Surface Plasmon Resonance (SPR): Immobilize the protein on a sensor chip and flow c-di-GMP over it. The change in refractive index provides real-time data on association and dissociation rates.
- In Vivo Functional Validation: Introduce point mutations in the identified binding residues (e.g., in the RXXD motif) into the P. aeruginosa chromosome. Compare Pel production and biofilm formation between the wild-type and mutant strains to confirm the functional importance of the binding site [32].
Protocol 3: Visualizing Exopolysaccharide in Biofilms
- Objective: To spatially localize specific exopolysaccharides (e.g., Psl) during biofilm development.
- Methodology:
- Biofilm Growth: Grow biofilms in flow cells or on submerged surfaces for varying durations.
- Fluorescent Staining: Use specific probes without disrupting the biofilm structure.
- For Psl: Use fluorescently conjugated lectins that specifically bind Psl monosaccharides (e.g., FITC-MOA for galactose, FITC-HHA for mannose) [28].
- For eDNA: Use stains like SYTO or TOTO.
- Confocal Laser Scanning Microscopy (CLSM): Image the stained biofilm to obtain high-resolution Z-stacks.
- Image Analysis: Use software (e.g., ImageJ, COMSTAT) to analyze the 3D architecture and quantify the spatial distribution and biomass of the polysaccharide matrix [28].
The workflow for a comprehensive study integrating these techniques is visualized below.
Figure 3: Experimental Workflow for EPS Regulation Studies. A multi-faceted approach combining genetics, phenotyping, microscopy, and biochemistry is essential to fully elucidate the role of a specific protein or signal in the EPS regulatory network.
The Scientist's Toolkit: Key Research Reagents and Materials
Advancing research in this field relies on a suite of specialized reagents and tools.
Table 4: Essential Research Reagent Solutions for EPS and Signaling Studies
| Reagent / Material | Function / Application | Specific Example / Note |
|---|---|---|
| c-di-GMP Analytical Standard | Calibration standard for LC-MS/MS quantification of cellular c-di-GMP levels. | Commercially available (e.g., BioLog). Essential for accurate absolute quantification. |
| Fluorescent Lectins (MOA, HHA) | Specific probes for visualizing Psl exopolysaccharide in biofilms via CLSM. | FITC-MOA binds galactose in Psl; FITC-HHA binds mannose [28]. |
| c-di-GMP Biosensors | Report intracellular c-di-GMP levels in vivo. | Plasmid-based systems expressing a c-di-GMP-responsive promoter fused to GFP. |
| Defined Mutant Libraries | Systematically study the function of genes encoding DGCs/PDEs and EPS biosynthesis. | Single-gene knockout mutants in PAO1 or PA14 background (e.g., PAO1 Transposon Mutant Library). |
| Anti-Psl Antibodies | Specific detection and quantification of Psl polysaccharide. | Used in ELISA, Western Blot, or for immunostaining as an alternative to lectins. |
| C-di-GMP Analogues | Tool for acutely manipulating intracellular c-di-GMP levels in experiments. | Membrane-permeable analogues (e.g., acetoxymethyl esters) can be used to raise levels. |
The regulatory network controlling EPS production in P. aeruginosa is a paradigm of bacterial complexity and adaptability. The central role of c-di-GMP in post-translationally activating machinery for Pel and Psl production, combined with its integration with the population-wide coordination of QS and other sensory inputs, creates a robust system that ensures biofilm formation occurs under favorable conditions. The specific focus on alginate, Psl, and Pel within this network highlights the multi-layered strategy P. aeruginosa employs: alginate for protective encapsulation, Psl for initial attachment and structural scaffolding, and Pel for structural integrity and niche-specific colonization.
Understanding these mechanisms at a deep level provides a roadmap for novel therapeutic interventions. Instead of targeting bacterial viability, which drives antibiotic resistance, future anti-biofilm strategies could aim to disrupt this regulatory network. Potential approaches include:
- Developing small-molecule inhibitors that block the catalytic sites of DGCs or the c-di-GMP binding pockets of receptors like PelD.
- Using peptide-based sequestrants that bind and neutralize intracellular c-di-GMP [30].
- Designing QS inhibitors (quorum quenching) to disrupt the coordinated expression of biofilm genes.
The integrated models, quantitative data, and experimental protocols outlined in this review provide a foundation for researchers and drug development professionals to explore these and other innovative strategies to combat persistent P. aeruginosa infections.
From Bench to Bedside: Techniques for EPS Analysis and Emerging Therapeutic Applications
In the study of Pseudomonas aeruginosa biofilms, the extracellular matrix is a defining feature, providing structural integrity and protection to the microbial community. This matrix is composed of a complex mixture of extracellular DNA, proteins, and notably, three self-secreted exopolysaccharides (EPSs)—alginate, Pel, and Psl. These EPSs are crucial for biofilm development, stability, and the increased tolerance to antibiotics observed in biofilm-associated infections, such as those in the cystic fibrosis (CF) lung [16]. Understanding the distinct roles and structural properties of each polysaccharide is paramount for developing effective anti-biofilm strategies. This guide details the core analytical techniques—lectin staining, confocal laser scanning microscopy (CLSM), mutant analysis, and structural elucidation methods—that enable researchers to dissect the composition, architecture, and function of the P. aeruginosa biofilm matrix.
Pseudomonas aeruginosa produces three primary exopolysaccharides, each with a unique chemical structure and functional role within the biofilm (Table 1).
Table 1: Characteristics of Major P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Primary Function in Biofilms | Biosynthetic Locus |
|---|---|---|---|
| Alginate | A high molecular weight, acidic polymer of non-repeating D-mannuronic acid and L-guluronic acid residues [4]. | Often the predominant EPS in mucoid strains from CF chronic infections; provides a viscous, protective gel that aids in immune evasion [16] [4]. | alg genes |
| Psl | A neutral, branched pentasaccharide composed of repeating D-mannose, D-glucose, and L-rhamnose subunits [17] [4]. | Critical for initial surface attachment and cell-cell interactions in non-mucoid strains; provides structural scaffolding and limits antibiotic penetration [17] [16]. | psl operon (15 genes, pslA-O) [16] |
| Pel | A cationic polymer of partially de-N-acetylated galactosamine and N-acetyl galactosamine residues [16]. | Important for pellicle formation and cell-to-cell interactions; can cross-link with eDNA to stabilize the matrix and sequester positively charged antibiotics like tobramycin [17] [16]. | pel operon (7 genes, pelA-G) [17] [16] |
The reliance on Pel or Psl as the primary structural polysaccharide is highly strain-dependent. For instance, the common laboratory strain PAO1 relies primarily on Psl, whereas the strain PA14 uses Pel, a difference attributed to a three-gene deletion in the psl operon of PA14 [17]. However, functional redundancy exists, as strains can evolve to upregulate one EPS to compensate for the loss of the other, ensuring biofilm integrity is maintained [17].
Analytical Techniques and Their Applications
Mutant Analysis
Purpose: Mutant analysis is a foundational genetic approach used to determine the specific contribution of each exopolysaccharide to biofilm formation, attachment, and structure.
Detailed Methodology:
- Strain Selection and Mutant Construction: Select P. aeruginosa strains of interest (e.g., clinical isolates, laboratory strains like PAO1 or PA14). Generate in-frame deletion mutants of key biosynthetic genes using standard allelic replacement strategies. Key target genes include:
pslD: A gene essential for Psl production. ApslDmutant is deficient in Psl biosynthesis and typically shows impaired attachment [17].pelF: A gene essential for Pel production. ApelFmutant is deficient in Pel biosynthesis [17].- Double
pelF pslDmutant: To investigate functional redundancy and the complete loss of both structural polysaccharides.
- Phenotypic Assays:
- Growth Curve Analysis: Confirm that any observed phenotypic changes are not due to general growth defects by comparing the growth rates of mutant and parent strains in planktonic culture (e.g., in rich medium like LB) [17].
- Initial Attachment Assay (Microtiter Dish Assay):
- Grow cultures statically in a microtiter dish for a short period (e.g., a few hours).
- Gently wash away non-adherent cells.
- Stain adherent cells with crystal violet, elute the dye, and measure the optical absorbance to quantify attachment biomass [17].
- Mature Biofilm Assay: Perform the microtiter dish assay with an extended incubation period (e.g., 24-48 hours) to assess the role of EPS in mature biofilm structure [17].
- Data Interpretation: Mutational analysis has demonstrated that Psl plays a critical role in surface attachment for most tested strains, as
pslmutants are consistently deficient in this initial step. In contrast, the role of Pel is more variable and strain-dependent, often becoming significant in the later stages of biofilm development and microcolony formation [17].
Lectin Staining and Fluorescent Labeling
Purpose: Lectin staining allows for the specific detection and spatial visualization of exopolysaccharides within the biofilm matrix, leveraging the binding specificity of lectins to sugar monomers.
Detailed Methodology:
- Lectin Selection: Choose fluorescently conjugated lectins based on the polysaccharide target (Table 2).
- Biofilm Preparation and Fixation: Grow biofilms on a suitable substrate (e.g., glass coverslip, ASEM dish). Gently rinse to remove non-adherent cells and fix with a mild fixative like paraformaldehyde.
- Staining Protocol: Incubate the fixed biofilm with the fluorescent lectin solution. For multiplex labeling of different species or components, other fluorescent dyes like CellTrace can be used simultaneously [34].
- Imaging and Analysis: Visualize using Confocal Laser Scanning Microscopy (CLSM). Acquire Z-stacks and use image analysis software to create 3D reconstructions and quantify biofilm parameters like biovolume and thickness.
Table 2: Research Reagent Solutions for Lectin Staining
| Reagent | Target Polysaccharide / Application | Specificity and Function |
|---|---|---|
| Wheat Germ Agglutinin (WGA) | Psl [35] [4] | Binds to N-acetylglucosamine and sialic acid residues; used with colloidal gold or fluorescent conjugates (e.g., Alexa 488) to label Psl [35]. |
| CellTrace Fluorescent Dyes (e.g., Far Red, Yellow, CFSE) | Multiplex species labeling in mixed biofilms [34] | Cell-permeant dyes that stably label the cytoplasm of live bacteria, enabling tracking of up to four different species in a single biofilm for up to 4 days via CSDM [34]. |
| CellMask Orange | General polysaccharide / matrix staining [4] | A lipophilic dye that incorporates into membranes and can be used to counterstain polysaccharide matrices in biofilms for CLSM visualization [4]. |
Confocal Laser Scanning Microscopy (CLSM)
Purpose: CLSM provides high-resolution, three-dimensional imaging of live, hydrated biofilms, enabling the analysis of biofilm architecture, EPS distribution, and cellular localization without destructive sample preparation.
Detailed Methodology:
- Biofilm Cultivation: Grow biofilms in flow-cell reactors or on other optically transparent surfaces that allow for continuous nutrient supply and waste removal, mimicking more natural biofilm growth conditions [17].
- Fluorescent Tagging: Express fluorescent proteins (e.g., GFP) in the bacterial cells. Use lectin staining (as above) or other fluorescent probes to label the extracellular matrix components.
- Image Acquisition: Use a CLSM to collect optical sections (Z-stacks) through the depth of the biofilm. The high-resolution capability of CLSM can reveal the fabric-like matrix of Psl connecting cells or the failure of mutant strains (e.g.,
pslorpelmutants) to develop beyond a monolayer [17] [4]. - Image Analysis: Employ software to reconstruct 3D models from Z-stacks. This allows for the quantification of biofilm biomass, volume, and substratum coverage. CLSM has been instrumental in showing that Psl forms a connecting matrix between cells and that Pel is crucial for the structural integrity of PA14 biofilms [4].
Structural Elucidation Methods
Purpose: Determining the precise chemical structure of exopolysaccharides is fundamental to understanding their physical properties and functional roles.
Detailed Methodology:
- Polysaccharide Purification: Isolate and purify the EPS from bacterial culture supernatants using methods such as ethanol precipitation and chromatographic techniques.
- Compositional Analysis:
- Use chemical and enzymatic hydrolysis to break down the polymer into its constituent monosaccharides.
- Analyze the hydrolysate using techniques like high-performance anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD) or gas chromatography-mass spectrometry (GC-MS) to identify and quantify the sugar monomers.
- Determination of Linkage and Sequence:
- Advanced Microscopy for Native Structure:
- Atmospheric Scanning Electron Microscopy (ASEM): This technique allows for the nanoscale visualization of delicate biofilms immersed in aqueous solution. Biofilms are cultured on an electron-transparent film and imaged from below with an inverted SEM, revealing native structures like fibrils, membrane vesicles, and water channels without the artifacts induced by air-drying [35].
- Atomic Force Microscopy (AFM): AFM can probe the surface of hydrated biofilms, providing information on the physical properties of the matrix. For example, it has shown alginate to be a soft, loosely adhered polymer surrounding the cells [4].
Experimental Workflow and Signaling Pathways
The investigation of biofilm exopolysaccharides follows a logical progression from genetic manipulation to structural analysis, with regulatory pathways governing EPS production. The following diagram outlines the core experimental workflow.
Figure 1: Core analytical workflow for EPS analysis.
The expression of the pel and psl operons is tightly regulated by a complex network of intracellular signaling molecules and transcriptional regulators, as illustrated below.
Figure 2: Key regulatory pathways for Pel and Psl synthesis.
The study of biofilms is critical for understanding chronic bacterial infections and developing effective treatments. For the opportunistic pathogen Pseudomonas aeruginosa, biofilm formation is a key virulence factor, heavily dependent on a matrix of self-secreted exopolysaccharides—alginate, Psl, and Pel [16]. These polymers provide structural integrity and protection, making biofilms highly tolerant to antibiotics and host immune responses [16]. This technical guide details the primary model systems—microtiter plates, flow cells, and animal models—used to dissect the roles of these exopolysaccharides in biofilm biology, providing researchers with robust methodologies for their investigations.
The Exopolysaccharide Matrix ofP. aeruginosa
The functional redundancy and unique roles of alginate, Psl, and Pel create a robust scaffold for P. aeruginosa biofilms. Table 1 summarizes their distinct characteristics and functions.
Table 1: Key Exopolysaccharides in the P. aeruginosa Biofilm Matrix
| Exopolysaccharide | Primary Monosaccharide Composition | Key Genetic Locus | Major Function in Biofilms |
|---|---|---|---|
| Alginate | Uronic acids (D-mannuronate and L-guluronate) [7] | alg (e.g., algD operon) | Overproduced in mucoid strains; confers resistance to antibiotics and host defenses; associated with chronic CF lung infection [16] [7]. |
| Psl | D-Mannose, D-Glucose, L-Rhamnose (neutral pentasaccharide) [16] | pslA-O | Critical for initial surface attachment and cell-cell interactions; provides structural integrity in mature biofilms; promotes tolerance to aminoglycosides [36] [16]. |
| Pel | Partially de-N-acetylated N-acetylgalactosamine and galactosamine (cationic polymer) [16] | pelA-G | Essential for pellicle formation; provides structural redundancy with Psl; cross-links with eDNA; sequesters positively charged antibiotics like tobramycin [36] [16]. |
The production of these exopolysaccharides is not mutually exclusive. Strains can produce one, two, or all three, and they exhibit significant functional redundancy. For instance, if the primary structural polysaccharide is lost via mutation, strains can acquire compensatory mutations that upregulate another, thereby preserving the capacity to form a biofilm [36] [17]. This redundancy underscores the need for model systems that can accurately probe the complex and dynamic interactions within the biofilm matrix.
Microtiter Plate Biofilm Assays
The microtiter plate (or microtiter dish) assay is a cornerstone of biofilm research due to its high-throughput capability, simplicity, and low cost. It is particularly effective for studying the early stages of biofilm formation, including initial attachment, and for genetic screens or testing biofilm formation under various conditions [37].
Experimental Protocol: Crystal Violet Biofilm Assay
The following protocol, adapted from O'Toole (2011), is a standard method for quantifying biofilm biomass in a 96-well plate [37].
1. Biofilm Growth:
- Grow an overnight culture of P. aeruginosa in a rich medium like LB.
- Dilute the culture 1:100 into a fresh, biofilm-promoting medium. For P. aeruginosa, a common medium is M63 minimal medium supplemented with 1 mM MgSO₄, 0.2% glucose, and 0.5% casamino acids. Using arginine as a sole carbon source can stimulate a more robust biofilm with less planktonic growth [37].
- Dispense 100 µL of the diluted culture per well into a sterile, flat-bottomed 96-well polystyrene microtiter plate. Include wells with sterile medium as negative controls.
- Incubate the plate statically for 4-24 hours at 37°C. The optimal incubation time should be determined empirically for each strain.
2. Biofilm Staining:
- After incubation, carefully invert the plate to dump out the planktonic cells and media.
- Rinse the plate by gently submerging it in a tub of water to remove loosely attached cells. Shake out the water and repeat this wash step a second time. Blot the plate vigorously on paper towels.
- Add 125 µL of a 0.1% (w/v) crystal violet (CV) solution to each well. Incubate at room temperature for 10-15 minutes.
- Dump out the CV and rinse the plate 3-4 times with water as before, ensuring all unbound dye is removed. Blot and air-dry the plate completely.
3. Biofilm Quantification:
- Add 125 µL of 30% acetic acid (in water) to each well to solubilize the CV bound to the biofilm.
- Incubate for 10-15 minutes at room temperature.
- Transfer 125 µL of the solubilized CV solution to a new flat-bottomed microtiter plate.
- Measure the absorbance at 550 nm using a plate reader, with 30% acetic acid as a blank [37].
Data Interpretation and Key Considerations
This assay quantifies total biofilm biomass. For example, it can clearly demonstrate that a psl mutant of strain PAO1 is severely defective in initial surface attachment, while a pel mutant is not [17]. Conversely, in a PA14 background, a pel mutation leads to a significant reduction in mature biofilm biomass [17].
Interlaboratory studies have shown that while the CV assay is versatile, its reproducibility can be enhanced with careful experimental design. The reproducibility standard deviation (SR) for CV in control experiments has been reported to be 0.44 on the log10 scale [38]. For higher reliability in antimicrobial efficacy testing, plate counts can be more responsive and reproducible (Slope/SR = 1.02) [38].
Table 2: Research Reagent Solutions for Microtiter Plate Assays
| Reagent / Material | Function / Application | Example Specification |
|---|---|---|
| Polystyrene 96-Well Plate | Substrate for static biofilm growth. | Flat-bottomed, untreated [38]. |
| Crystal Violet (0.1%) | Dye that stains biofilm biomass (e.g., polysaccharides, proteins). | Aqueous solution; handle with care as it stains [37]. |
| 30% Acetic Acid | Solubilizes crystal violet dye for spectrophotometric quantification. | In water; used as blank and elution solution [37]. |
| M63 Minimal Medium | Defined medium for controlled, robust biofilm formation. | Supplemented with MgSO₄, glucose, and casamino acids [37]. |
| Resazurin Stain | Metabolic stain to assess viable cell activity in biofilms. | Fluorescence measurement (λex 560 nm / λem 590 nm) [38]. |
Flow Cell Biofilm Models for Architectural Analysis
While microtiter plates are excellent for biomass quantification, flow cell reactors are the system of choice for studying the dynamic, three-dimensional architecture of mature biofilms under conditions that mimic flowing environments, such as those found in catheters or the CF lung.
Flow cells consist of small chambers with a cover glass substratum through which fresh medium is continuously perfused. This setup allows for the real-time, non-destructive observation of biofilm development using techniques like confocal laser scanning microscopy (CLSM).
Key Protocol Steps:
- Assembly and Sterilization: The flow cell system is assembled and sterilized, often by flowing a sterilizing solution like sodium hypochlorite followed by sterile water.
- Inoculation: The system is inoculated with a bacterial suspension, which is allowed to attach to the cover glass surface during a static phase.
- Continuous Flow: Fresh, dilute medium is pumped through the system at a constant, low rate (e.g., 0.2 mL/min) for several days to support biofilm development without promoting excessive planktonic growth.
- Staining and Imaging: At desired time points, biofilms can be stained with fluorescent dyes specific for cells (e.g., SYTO dyes) and matrix components (e.g., lectins that bind specific exopolysaccharides like Psl) and visualized using CLSM [17].
Elucidating Exopolysaccharide-Specific Roles
Flow cell studies have been instrumental in defining the distinct architectural contributions of Pel, Psl, and alginate:
- Psl-Dependent Architecture: The common lab strain PAO1 primarily utilizes Psl and forms characteristic mushroom-shaped microcolonies in flow cells. A psl mutant in this strain is arrested at the monolayer stage, failing to develop these complex structures [17].
- Pel-Dependent Architecture: Strain PA14, which has a deletion in the psl operon, relies on Pel for mature biofilm formation. A PA14 pel mutant similarly fails to progress beyond a monolayer in a flow cell [17].
- Alginate's Role: Alginate-overproducing mucoid strains, common in CF, form large, finger-like microcolonies. Alginate also contributes to cell viability within the biofilm structure [7].
- Complex Interactions: Mutants lacking multiple exopolysaccharides reveal interactive roles. For instance, a mutant producing only alginate (lacking both Psl and Pel) loses the ability to form biofilms altogether, while the loss of Psl can enhance Pel production, and vice versa, demonstrating compensatory regulation [7].
Animal Models for ComplexIn VivoStudy
Animal models provide the necessary complexity to study biofilm infections within a living host, encompassing the dynamics of the immune system, nutrient gradients, and tissue pathology. They are indispensable for validating findings from in vitro systems and for preclinical testing of new therapeutics.
Commonly Utilized Infection Models
Table 3 outlines key animal models used to study P. aeruginosa biofilm infections relevant to human disease.
Table 3: Animal Models for P. aeruginosa Biofilm Infection Studies
| Infection Type | Exemplified Human Disease | Common Animal Model & Method | Key Application in Biofilm/Exopolysaccharide Research |
|---|---|---|---|
| Chronic Respiratory | Cystic Fibrosis (CF) Lung Infection | Transgenic CFTR-/- mice (e.g., Cftrtm1UNC); intratracheal instillation [39]. | Studies on alginate-overproducing mucoid variants and chronic, antibiotic-tolerant infections [39]. |
| Wound Infection | Diabetic Foot Ulcers (DFU), Burn Wounds | Murine full-thickness excision or burn wound; topical application of bacteria [39] [40]. | Modeling polymicrobial biofilms; evaluating anti-biofilm therapies like phage cocktails targeting Psl/Pel [40]. |
| Acute Systemic | Sepsis, Bacteremia | Mouse intravenous or intraperitoneal injection [39]. | Assessing the role of exopolysaccharides in systemic virulence and survival. |
| Keratitis | Contact Lens-Associated Keratitis | Mouse cornea; scratch and inoculation model [39]. | Investigating host-pathogen interactions in a localized tissue infection. |
Insights from Animal Models and Advanced 3D Systems
Animal models have confirmed the critical role of biofilms in chronic infections. For example, in CF lung models, the biofilm matrix, rich in alginate, Psl, and Pel, contributes significantly to the failure of antibiotic therapy [16]. Furthermore, the persistence of biofilms in diabetic wound models highlights the protective nature of the matrix against host immune cells [40].
To bridge the gap between traditional in vitro models and complex animal studies, advanced 3D bioengineered models are being developed. These include:
- 3D Bioengineered Skin: Constructs using biomaterials (e.g., collagen, hydrogels) seeded with fibroblasts and keratinocytes to mimic human skin, including diabetic skin conditions [40].
- Hydrogel-based Systems: Alginate or other polymer hydrogels that create a 3D microenvironment to grow biofilms, better simulating the tissue-embedded nature of infections like DFUs [40].
These 3D models aim to more accurately replicate the host microenvironment—including factors like hyperglycemia, hypoxia, and polymicrobial communities—for high-quality, high-throughput testing of novel anti-biofilm strategies, such as phage therapy, while reducing animal use [40].
The strategic use of microtiter plates, flow cells, and animal models provides a complementary toolkit for deconstructing the complex, exopolysaccharide-driven biology of P. aeruginosa biofilms. Microtiter plates offer unmatched throughput for screening and initial characterization, flow cells reveal the exquisite architecture of mature biofilms, and animal models contextualize these findings within the multifaceted environment of a living host. The continued refinement of these models, including the development of sophisticated 3D systems, is paramount for advancing our understanding of biofilm-mediated pathogenesis and for translating this knowledge into effective clinical interventions against resilient biofilm infections.
The biofilm mode of growth is a critical virulence factor for Pseudomonas aeruginosa, particularly in chronic infections such as those in the cystic fibrosis (CF) lung. This resilience is largely conferred by the extracellular polymeric substance (EPS), a matrix that provides structural integrity and protects embedded cells from antibiotics and host immune responses [41] [16]. The three self-secreted exopolysaccharides—alginate, Psl, and Pel—are key components of this matrix, each contributing uniquely to biofilm development, stability, and pathogenicity [42] [43]. Alginate is characteristic of mucoid strains prevalent in CF and provides a protective barrier [41]. Psl is critical for surface attachment and initial structural scaffolding, while Pel provides cationic cross-linking and enhances tolerance to specific antibiotics like aminoglycosides [44] [16]. The inherent redundancy between Pel and Psl further complicates treatment, as strains can compensate for the loss of one polysaccharide by upregulating the other [45] [46]. Enzymatic degradation of these EPS components has emerged as a promising therapeutic strategy to disrupt biofilms, resensitize bacteria to conventional antibiotics, and improve clinical outcomes. This technical guide details the current knowledge and methodologies for using alginate lyase and Psl-degrading enzymes as anti-biofilm agents.
Target Exopolysaccharides: Structure and Function
A thorough understanding of the target exopolysaccharides is a prerequisite for developing effective enzymatic strategies.
Table 1: Characteristics of Key P. aeruginosa Exopolysaccharides
| Polysaccharide | Chemical Structure | Primary Role in Biofilm | Impact on Antibiotic Efficacy |
|---|---|---|---|
| Alginate | Linear copolymer of β-D-mannuronic acid and α-L-guluronic acid [41]. Heavily acetylated [41]. | Forms a protective, mucoid capsule; key in chronic CF infections [41] [16]. | Limits antibiotic penetration; provides a physical diffusion barrier [16]. |
| Psl | Neutral, branched pentasaccharide of D-mannose, D-glucose, and L-rhamnose [16]. | Primary scaffold in many strains; essential for initial surface attachment and cell-cell adhesion [42] [16]. | Sequesters antibiotics; prevents complement deposition and neutrophil phagocytosis [16]. |
| Pel | Cationic polymer of partially de-N-acetylated galactosamine and N-acetylgalactosamine [16]. | Provides structural redundancy with Psl; crucial for cell-cell adhesion and pellicle formation [44] [45]. | Confers specific tolerance to aminoglycoside antibiotics via cationic sequestration [44] [16]. |
The functional interplay between these polymers is complex. Alginate production is associated with chronic, mucoid infections and provides a general physical barrier [41]. In contrast, Psl and Pel often serve overlapping structural roles as primary scaffold components in non-mucoid biofilms, with their individual importance varying significantly between strains [45]. Pel's unique cationic nature allows it to cross-link with negatively charged extracellular DNA (eDNA), further reinforcing the matrix and protecting the eDNA from degradation by nucleases [16]. The regulatory network controlling the expression of these EPS is sophisticated, with evidence of inverse regulation; for example, a lack of Pel can enhance alginate production and vice versa [42].
Visualizing EPS-Specific Anti-Biofilm Enzyme Mechanisms
The following diagram illustrates the targeted mechanism of action for alginate lyase and the Psl-degrading enzyme, PslG, against the structural components of the P. aeruginosa biofilm matrix.
Enzyme Classes and Their Experimental Applications
Alginate Lyases
Alginate lyases (EC 4.2.2.3) are enzymes that catalyze the β-elimination of glycosidic bonds in alginate, producing unsaturated oligosaccharides. They are categorized based on their cleavage pattern (endo- vs. exo-lytic) and substrate preference (poly-M, poly-G, or both) [47].
Table 2: Characterized Alginate Lyases for Anti-Biofilm Applications
| Enzyme Name | Source Organism | Type | Optimal Activity Conditions | Key Experimental Findings |
|---|---|---|---|---|
| CaAly | Cellulophaga algicola [47] | Endolytic [47] | Not fully specified [47] | Superior biofilm inhibition against a mucoid CF clinical isolate (P. aeruginosa 2843) [47]. |
| VspAlyVI | Vibrio sp. QY101 [47] | Endolytic [47] | 40°C, pH 7.5 (activity enhanced by Zn²⁺) [47] | Effective biofilm inhibition against a mucoid CF clinical isolate [47]. |
| FspAlyFRB | Falsirhodobacter sp. alg1 [47] | Exolytic [47] | 25-30°C, pH 6-9 [47] | Effective biofilm inhibition against a mucoid CF clinical isolate [47]. |
| SA1-IV | Sphingomonas sp. A1 [47] | Exolytic [47] | N/D | Effective biofilm inhibition against a mucoid CF clinical isolate [47]. |
| PspAlgL | Pseudomonas sp. QD03 [47] | Endolytic [47] | 37°C, pH 7.5 (enhanced by Ca²⁺, not inhibited by Zn²⁺) [47] | Active against acetylated alginate; ideal for CF sputum with high divalent cation content [47]. |
Psl-Degrading Enzymes
The glycoside hydrolase PslG has shown remarkable efficacy in targeting the Psl polysaccharide. PslG catalyzes the hydrolysis of glycosidic bonds within the Psl pentasaccharide repeats. Studies demonstrate that even sub-inhibitory concentrations of PslG can disrupt biofilm structure and potentiate the effect of antibiotics like tobramycin [16]. Notably, the catalytic activity of PslG is primarily localized to its N-terminal domain, and the enzyme is thought to act on the Psl polysaccharide directly associated with the bacterial cell surface, causing a rapid disintegration of the biofilm matrix [47] [16].
Protocol: Assessing Anti-Biofilm Efficacy of Enzymes In Vitro
The following is a standard experimental protocol for evaluating the efficacy of alginate lyases or PslG in inhibiting biofilm formation, adapted from published methodologies [47].
Objective: To quantify the ability of purified enzymes to prevent biofilm formation by a mucoid P. aeruginosa strain.
Materials:
- Bacterial Strain: Mucoid P. aeruginosa (e.g., clinical isolate from CF sputum) [47].
- Growth Medium: Cation-adjusted Mueller Hinton Broth (CAMHB) [47].
- Enzymes: Purified recombinant alginate lyase (e.g., CaAly) or PslG with a C-terminal 6xHis tag [47].
- Equipment: 96-well flat-bottom microtiter plate, sterile incubator, microplate reader.
- Reagents: Methanol, 0.1% (w/v) crystal violet solution, acetic acid (33% v/v).
Method:
- Protein Purification: Clone the codon-optimized gene for the target enzyme (e.g., pslG or alginate lyase) into an expression vector like pET-28a(+). Express the recombinant protein in E. coli BL21(DE3) with IPTG induction. Purify the 6xHis-tagged protein from the clarified cell lysate using Ni-NTA affinity chromatography. Dialyze the eluted protein extensively against Tris-buffered saline (TBS: 20 mM Tris, 150 mM NaCl, pH 7.5) to remove imidazole. Determine protein concentration and purity via SDS-PAGE and spectrophotometry [47].
- Biofilm Assay:
- Prepare an overnight culture of the mucoid P. aeruginosa strain in CAMHB.
- Dilute the culture to a density of approximately 1 x 10⁷ CFU/mL in fresh CAMHB.
- Dispense 200 µL of the bacterial suspension into the wells of a 96-well microtiter plate.
- Add the purified enzyme to test wells at desired final concentrations (e.g., 0.1-10 µg/mL). Include control wells with CAMHB only (negative control) and bacteria without enzyme (positive control). Perform all treatments in triplicate.
- Incubate the plate statically at 37°C for 24 hours.
- Biofilm Quantification (Crystal Violet Staining):
- Carefully remove the planktonic cells and medium from the wells.
- Fix the adherent biofilm cells by adding 200 µL of methanol to each well for 15 minutes.
- Discard methanol and allow the plate to air dry.
- Stain the biofilm with 200 µL of 0.1% crystal violet solution for 15-20 minutes.
- Gently rinse the plate under running tap water to remove excess stain.
- Elute the bound crystal violet by adding 200 µL of 33% acetic acid to each well.
- Measure the absorbance of the eluted dye at 550-600 nm using a microplate reader.
- Data Analysis: Compare the average absorbance of the enzyme-treated wells to the untreated positive control. The percentage of biofilm inhibition is calculated as:
[1 - (Abs_treated / Abs_control)] * 100.
Protocol: Biofilm Penetration Assay Using Enzyme-Functionalized Microrobots
An emerging application for enzymes is their integration into advanced drug delivery systems. The following protocol details a novel method for constructing and testing enzyme-coated microrobots for enhanced biofilm penetration [48].
Objective: To fabricate self-propelled biohybrid microrobots capable of degrading and penetrating P. aeruginosa biofilms.
Materials:
- Microalgae: Chlamydomonas reinhardtii (CR) [48].
- Enzyme: Alginate lyase (Alg) [48].
- Chemical Reagents: Bio-orthogonal reaction reagents (e.g., for click chemistry) [48].
- Equipment: Confocal laser scanning microscope (CLSM), flow cell system for biofilm growth.
Method:
- Microrobot Fabrication (CR@Alg):
- Harvest and wash Chlamydomonas reinhardtii microalgae to remove culture media.
- Functionalize the surface of the microalgae with alginate lyase via a bio-orthogonal reaction (e.g., strain-promoted azide-alkyne cycloaddition). This typically involves pre-modifying the algal surface and the enzyme with complementary reactive groups.
- Purify the resulting CR@Alg microrobots to remove unbound enzyme.
- Biofilm Growth and Imaging:
- Grow a mature P. aeruginosa biofilm (e.g., a mucoid strain) in a flow cell system for 3-5 days to establish a robust 3D structure.
- Treat the biofilm by introducing a suspension of CR@Alg microrobots.
- Penetration and Efficacy Analysis:
- Use live-cell imaging and CLSM to track the movement and penetration depth of the fluorescently labeled microrobots into the biofilm over time. The study cited reported a 2.4-fold increase in penetration compared to unmodified microalgae [48].
- To assess therapeutic efficacy, after allowing the microrobots to penetrate, expose the biofilm to laser irradiation at an appropriate wavelength. The chlorophyll within the microalgae will act as an endogenous photosensitizer, generating reactive oxygen species (ROS) and triggering photodynamic therapy (PDT) [48].
- Quantify bacterial killing using viable plate counts or live/dead bacterial staining kits.
The workflow for this advanced experimental approach is visualized below.
The Scientist's Toolkit: Key Research Reagents and Solutions
Table 3: Essential Reagents for Enzymatic Anti-Biofilm Research
| Reagent / Material | Specifications / Key Features | Primary Function in Research |
|---|---|---|
| Mucoid P. aeruginosa Strain | Clinical isolate from cystic fibrosis sputum (e.g., strain 2843) [47]. Alginate-overproducing phenotype. | Physiologically relevant model for testing anti-biofilm therapies against chronic infections. |
| Cation-Adjusted Mueller Hinton Broth (CAMHB) | Standardized for antimicrobial susceptibility testing; cation concentration adjusted per CLSI guidelines [47]. | Culture medium for consistent and reproducible biofilm growth in microtiter assays. |
| pET-28a(+) Expression Vector | E. coli expression plasmid with T7 promoter and N-terminal His-Tag sequence [47]. | Cloning and high-yield recombinant expression of 6xHis-tagged alginate lyases and PslG. |
| Ni-NTA Affinity Resin | Nickel-charged resin for immobilized metal affinity chromatography (IMAC) [47]. | One-step purification of recombinant 6xHis-tagged enzymes from clarified cell lysates. |
| Crystal Violet Solution | 0.1% (w/v) in water [47]. | Histochemical stain for quantifying total adherent biofilm biomass after fixation. |
| Chlamydomonas reinhardtii | Wild-type, motile strain of green microalgae [48]. | Platform for creating self-propelled, enzyme-delivery biohybrid microrobots. |
Enzymes such as alginate lyases and PslG represent a potent and specific strategy to dismantle the protective EPS matrix of P. aeruginosa biofilms. The experimental data, including the ability of these enzymes to inhibit biofilm formation and their synergistic effects with traditional antibiotics, provide a compelling case for their development as therapeutic adjuvants. The innovation of coupling enzymes with delivery vehicles like self-propelling microrobots further enhances their potential by addressing the critical challenge of biofilm penetration [48]. However, significant hurdles remain for clinical translation. These include ensuring enzyme stability and activity in the harsh environment of the CF lung (e.g., in the presence of high divalent cation concentrations and proteases) [47], mitigating potential immunogenicity, and scaling up production. Future research must focus on robust in vivo efficacy studies, exploring combinations of enzymes that target multiple EPS components simultaneously (e.g., an alginate lyase + PslG cocktail), and engineering next-generation enzymes with enhanced catalytic properties and stability. The targeted degradation of the biofilm matrix continues to be a promising frontier in the fight against persistent P. aeruginosa infections.
The extracellular polymeric substance (EPS) matrix of bacterial biofilms represents a formidable barrier to effective antimicrobial therapy, contributing significantly to the persistence of chronic infections. This is particularly evident in Pseudomonas aeruginosa, an opportunistic pathogen notorious for its role in cystic fibrosis (CF) lung infections and hospital-acquired diseases. The biofilm matrix is a complex, hydrated polymer network primarily composed of polysaccharides, proteins, nucleic acids, and lipids that provides structural integrity and protection to encased microbial communities [49]. Within the context of P. aeruginosa, the exopolysaccharides alginate, Psl, and Pel play distinct yet complementary roles in biofilm architecture and function. Alginate, a high molecular weight, acetylated polymer of β-1,4 linked L-guluronic and D-mannuronic acids, predominates in mucoid strains frequently isolated from CF lungs [27]. Non-mucoid strains typically utilize Psl (a pentasaccharide of D-glucose, D-mannose, and L-rhamnose) and Pel (a cationic exopolysaccharide) for biofilm formation and stability [50] [27]. The anionic nature of these matrix components facilitates cation bridging, particularly with calcium ions (Ca²⁺), which cross-links the polymer network and enhances structural integrity [50] [51]. This review explores the strategic disruption of the EPS matrix as an adjuvant approach to potentiate conventional antimicrobials, with a specific focus on the molecular mechanisms and experimental methodologies driving this innovative therapeutic paradigm.
Mechanistic Foundations of EPS Disruption
Molecular Interactions and Disruption Strategies
The efficacy of EPS-targeting antibiotic adjuvants stems from their ability to interfere with critical molecular interactions within the biofilm matrix. Key mechanisms include disruption of cation bridging, direct enzymatic degradation of matrix components, and interference with matrix assembly and organization.
Disruption of Cationic Bridging: The anionic components of the EPS, particularly alginate and extracellular DNA (eDNA), form cationic bridges with Ca²⁺ ions that significantly contribute to matrix stability. Alginate oligosaccharides (OligoG CF-5/20), low molecular weight derivatives from marine algae, competitively chelate Ca²⁺ ions, disrupting the DNA-Ca²⁺-DNA bridges that reinforce the biofilm structure [50]. This disruption increases biofilm porosity and permeability, facilitating enhanced antibiotic diffusion. Isothermal titration calorimetry (ITC) and Fourier transform infrared (FTIR) spectroscopy have confirmed the direct interaction between OligoG and Ca²⁺, while molecular dynamics (MD) simulations provide atomistic details of these binding interactions [50].
Matrix Component Degradation and Structural Disassembly: Enzymatic degradation represents another strategic approach. Alginate lyase, for instance, directly cleaves alginate polymers, though its therapeutic efficacy has been limited, prompting advances in enzyme engineering [50]. Beyond polysaccharide targeting, disruption of the Psl matrix scaffolding has profound consequences. The Psl exopolysaccharide is anchored to the bacterial cell surface in a helical pattern and serves as a primary scaffolding component during early biofilm development [28]. Direct visualization using Psl-specific lectin staining (e.g., MOA, HHA) has demonstrated that chemical or enzymatic dissociation of Psl from the cell surface disrupts the entire matrix architecture, preventing microcolony formation and promoting biofilm dissociation [28].
Induction of Structural Heterogeneity and Dispersion: During biofilm maturation, programmed cell death and autolysis in the microcolony center create a cavity devoid of Psl matrix but enriched with eDNA and motile cells [28]. This structured cavity facilitates a critical life cycle event: seeding dispersal. Mutants with reduced autolysis cannot form this cavity, while those with elevated lysis form larger cavities and disperse prematurely [28]. This natural process highlights cell death and lysis as potential targets for adjuvants designed to induce biofilm dispersion, thereby sensitizing bacteria to antibiotics.
The diagram below synthesizes these core mechanisms into a coherent pathway linking adjuvant action to antibiotic potentiation.
Advanced Delivery Systems for Enhanced Efficacy
Overcoming the physical barrier of the EPS requires not only effective bioactive compounds but also advanced delivery systems that ensure their targeted release within the biofilm. Ultrasound-responsive nanoscale drug delivery platforms (nanodroplets) represent a cutting-edge approach. These phospholipid-coated nanodroplets can be loaded with antimicrobials and are designed to vaporize upon application of focused ultrasound (FUS). This vaporization achieves two critical objectives simultaneously: mechanical disruption of the biofilm structure and spatiotemporally controlled release of encapsulated agents [52]. Studies have demonstrated that this technology can dramatically reduce the required doses of antibiotics, achieving an average 7.6-fold reduction in the minimum bactericidal concentration (MBC) and a 44.4-fold reduction in the minimum biofilm eradication concentration (MBEC) compared to free drug treatment [52]. Furthermore, this platform alters the subcellular distribution of antimicrobials, enhancing their accumulation within the cytoplasm of biofilm-residing bacteria by 11.1-fold, thereby directly addressing the penetration and localization limitations that traditionally plague antibiofilm therapies [52].
Experimental Approaches for Evaluating Anti-Biofilm Adjuvants
Key Methodologies and Workflows
Rigorous evaluation of EPS-disrupting adjuvants relies on a combination of imaging, quantitative analysis, and molecular interaction studies. The following workflow outlines a standard integrated approach, from treatment to analysis.
Table 1: Core Quantitative Metrics for Anti-Biofilm Adjuvant Evaluation
| Metric | Experimental Method | Significance & Interpretation | Exemplar Data |
|---|---|---|---|
| Biomass Reduction | COMSTAT analysis of CLSM z-stacks | Indicates overall dissolution of the biofilm structure. | ≥0.5% OligoG: Significant reduction (P < 0.05) [50] |
| Biofilm Thickness | COMSTAT analysis of CLSM z-stacks | Measures the collapse of 3D biofilm architecture. | ≥2% OligoG: Significant decrease (P < 0.05) [50] |
| Roughness Coefficient | COMSTAT analysis of CLSM z-stacks | Reflects increased structural heterogeneity and disruption. | ≥6% OligoG: Significant increase (P < 0.05) [50] |
| Nanoparticle Diffusion | Fluorescent nanoparticle tracking | Quantifies increased matrix permeability and porosity. | OligoG treatment: Significant increase (P < 0.05) [50] |
| Antibiotic Potentiation | Minimum Biofilm Eradication Concentration (MBEC) | Measures the fold-reduction in antibiotic dose required to kill biofilm cells. | Ultrasound Nanodroplets: 44.4-fold avg. MBEC reduction [52] |
The Researcher's Toolkit: Essential Reagents and Materials
Table 2: Key Research Reagent Solutions for EPS Disruption Studies
| Reagent / Material | Function & Role in Experimentation | Specific Examples & Notes |
|---|---|---|
| Alginate Oligosaccharides (OligoG) | Low molecular weight adjuvant that chelates Ca²⁺, disrupting ionic bridges in the matrix. | OligoG CF-5/20 (Mn = 3200 g/mol, G-content >85%); used at 0.5%-6% (w/v) [50]. |
| Fluorescent Lectins | Specific labeling and visualization of exopolysaccharides in the biofilm matrix. | FITC-MOA & FITC-HHA for Psl; Concanavalin A (ConA) for levan [53] [28]. |
| COMSTAT Software | Image-analysis platform for quantifying 3D biofilm architecture from CLSM z-stack images. | Quantifies biomass, thickness, roughness, and surface-to-volume ratio [50]. |
| Isothermal Titration Calorimetry (ITC) | Directly measures thermodynamic parameters of molecular interactions (e.g., OligoG-Ca²⁺). | Confirms binding stoichiometry, affinity, and enthalpy changes [50]. |
| Ultrasound-Responsive Nanodroplets | Advanced delivery platform for spatiotemporal control of antimicrobial and adjuvant release. | Phospholipid-coated, perfluorocarbon-core; vaporize with FUS (~3.1 MPa) [52]. |
| Texas Red (TxRd) | Fluorescent dye for covalent labeling of molecules to track their diffusion and localization. | TxRd-OligoG conjugate confirmed oligomer penetration into established biofilms [50]. |
Quantitative Evidence and Therapeutic Application
Efficacy Data and Synergistic Potentiation
The therapeutic potential of EPS disruption is unequivocally demonstrated by quantitative data showing enhanced antibiotic efficacy against biofilm-encased bacteria. Treatment with alginate oligomers (OligoG ≥2%) significantly reduces the structural quantities of EPS polysaccharides and eDNA, which correlates with a marked improvement in antibiotic efficacy against established biofilms [50]. This synergistic relationship is not limited to conventional antibiotics but extends to novel therapeutic platforms. For instance, the integration of antimicrobials into ultrasound-triggered nanodroplets has shown a dramatic reduction in the concentrations required for eradication, with an average 25.5-fold reduction in the dose needed for complete persister cell eradication [52]. This underscores the dual benefit of matrix disruption: it not only improves antibiotic penetration but also appears to alter the physiological state of the bacteria, making them more susceptible to killing.
Table 3: Quantified Synergistic Effects of Combined Adjuvant-Antibiotic Treatments
| Adjuvant / Platform | Antibiotic / Challenge | Biofilm Model | Key Quantitative Outcome |
|---|---|---|---|
| OligoG (≥2%) | Colistin, others | Mucoid P. aeruginosa (NH57388A) | Significant potentiation of antibiotic activity (up to 512-fold in planktonic systems) [50]. |
| Ultrasound Nanodroplets (AZ-ND/FUS) | Azithromycin | Clinical MRSA & ESBL isolates | 13.45-fold reduction in metabolic MIC vs. free drug for E. coli [52]. |
| Ultrasound Nanodroplets (BF-ND/FUS) | Besifloxacin | Clinical MRSA & ESBL isolates | 15.72-fold reduction in MBC vs. free drug for E. coli [52]. |
| Psl-Disrupting Reagents | Not Applicable | P. aeruginosa PAO1 | Chemical dissociation of Psl disrupts biofilm structure on surfaces [28]. |
Clinical Translation and Future Perspectives
The transition of EPS-disrupting adjuvants from bench to bedside represents the next critical phase in combating biofilm-mediated resistance. The clinical imperative is clear: biofilms contribute to the recalcitrance of chronic lung infections in cystic fibrosis, non-healing wounds, and infections associated with medical devices [50] [54]. The ongoing clinical investigation of OligoG highlights the promising translational pathway for this class of agents [50]. Future development will likely focus on optimizing combination therapies that pair matrix-disrupting adjuvants with conventional antibiotics, as well as engineering sophisticated delivery systems like nanocarriers and engineered bacteriophages to ensure precise targeting and release within complex infection sites [54] [55] [52]. Furthermore, an 'omics'-driven discovery approach holds potential for identifying novel natural antibiotics and potentiators from diverse biological sources, which can be developed sustainably to bridge the antibiotic discovery-resistance gap [56]. While challenges in reproducibility, standardization, and scalability remain, the strategic disruption of the biofilm matrix undeniably offers a powerful avenue to restore the efficacy of existing antibiotics and address the global crisis of antimicrobial resistance.
Pseudomonas aeruginosa is a formidable opportunistic pathogen responsible for chronic infections in immunocompromised patients, particularly those with cystic fibrosis (CF), burn wounds, or undergoing medical device implantation. Its notorious persistence in clinical settings is primarily attributed to its ability to form antibiotic-resistant biofilms [25]. These biofilms are structured communities of bacterial cells encased in a self-generated matrix of extracellular polymeric substances (EPS) that function as a scaffold, protecting encased bacteria from environmental stresses, impeding phagocytosis, and conferring remarkable capacity for long-term colonization [25]. Biofilm-associated bacteria can exhibit tolerance to antimicrobial agents up to 1000 times greater than their planktonic counterparts [25].
The biofilm matrix of P. aeruginosa contains three self-secreted exopolysaccharides of paramount biomedical relevance: alginate, Psl, and Pel. Each plays a distinct but crucial role in biofilm integrity, mechanical stability, and antimicrobial defense [16]. Alginate, a negatively charged acetylated polymer of mannuronic and guluronic acids, is overproduced by mucoid strains predominant in chronic CF lung infections and helps in evading host immune responses [25] [16]. Psl, a neutral pentasaccharide, is critical for initial surface attachment, cell-to-cell interactions, and maintaining the structural stability of mature biofilms [25] [16]. Pel, a cationic polymer of partially deacetylated N-acetylgalactosamine, facilitates cell-to-cell interactions and provides specific protection against aminoglycoside antibiotics [57] [16]. Targeting these exopolysaccharides with degrading enzymes presents a promising therapeutic strategy to disrupt the biofilm physical barrier and potentiate conventional antibiotics. This whitepaper examines the formulation challenges and development pathways for these enzyme-based adjuvants.
Structural Foundations of theP. aeruginosaBiofilm Matrix
Understanding the distinct chemical and physical properties of each exopolysaccharide is fundamental to designing effective enzymatic countermeasures. The table below summarizes the key characteristics of the three primary exopolysaccharides.
Table 1: Key Characteristics of P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Charge | Primary Role in Biofilm | Role in Antibiotic Tolerance |
|---|---|---|---|---|
| Alginate | Polymer of β-D-mannuronate (M) and α-L-guluronate (G) [58] | Negative [6] | Maturation, protection from phagocytosis; major component in mucoid strains [25] | Reduces antibiotic diffusion; implicated in viscoelastic barrier formation [6] [16] |
| Psl | Pentasaccharide of D-glucose, D-mannose, and L-rhamnose [25] [16] | Neutral [6] | Initial surface attachment, cell-to-cell interactions, structural scaffold [25] [16] | Sequesters antibiotics; prevents neutrophil phagocytosis [25] [16] |
| Pel | Polymer of partially deacetylated N-acetyl-D-glucosamine and N-acetyl-D-galactosamine [9] [25] | Positive [6] | Cell-to-cell interactions, pellicle formation, biofilm integrity [57] [25] | Confers tolerance to aminoglycosides and colistin [57] [25] |
The architecture of the biofilm is not merely a static mixture of these components. Recent structural biology breakthroughs have revealed the sophisticated machinery required for polysaccharide secretion. For instance, the Pel export complex (PelBC) forms a 250 kDa structure where a periplasmic ring of twelve lipoprotein subunits (PelC) is mounted atop a nanodisc-embedded β-barrel (PelB) [9]. The negatively charged interior of the PelB β-barrel facilitates the transport of the cationic Pel polysaccharide, with molecular dynamics simulations suggesting a flexible loop (Plug-S) acts as a gate for export [9]. This level of detail is crucial for understanding biosynthesis and identifying potential inhibitory sites.
Enzyme-Based Therapeutic Strategies and Their Mechanisms of Action
Enzymatic degradation of the biofilm matrix is a precision approach to disrupt this physical barrier. Different enzymes target specific polysaccharides, and their efficacy is highly dependent on substrate specificity.
Table 2: Biofilm-Degrading Enzymes and Their Characteristics
| Enzyme | Target EPS | Polymer Substrate Specificity | Key Findings and Efficacy |
|---|---|---|---|
| Alg2A (PL7 family) | Alginate | PolyM/G (broad-spectrum) [58] | Effective at dissolving biofilms; synergistic effect with ciprofloxacin demonstrated [58] |
| A1-II' (PL7 family) | Alginate | PolyM/G (bifunctional) [58] | Effective at dissolving biofilms; requires dual polyM/polyG activity for efficacy [58] |
| AlyA1 (PL7 family) | Alginate | PolyG-specific [58] | Less effective at dissolving biofilms alone [58] |
| A1-III (PL5 family) | Alginate | PolyM-specific [58] | Less effective at dissolving biofilms alone [58] |
| Psl Glycoside Hydrolase | Psl | Psl polysaccharide [20] | Shown to inhibit biofilm formation on 3D lung epithelial cell aggregates [20] |
| Pel-Specific Hydrolase | Pel | Information not fully elucidated | Potential target, but enzymes are less characterized compared to alginate and Psl lyases. |
The therapeutic mechanism of these enzymes is twofold. First, they directly dismantle the structural scaffold of the biofilm, breaking down the physical barrier that impedes antibiotic penetration. Second, by disrupting the matrix, they expose the embedded bacterial cells to host immune defenses and increase their susceptibility to co-administered antimicrobials [16]. For example, enzymes with broad-spectrum activity against both polyM and polyG blocks of alginate (e.g., Alg2A) are significantly more effective than those with narrow specificity, highlighting the importance of enzyme selection [58].
Figure 1: Mechanism of Action for EPS-Degrading Enzymes as Antibiotic Adjuvants. The diagram illustrates the synergistic process where enzymatic disruption of the biofilm matrix enhances antibiotic efficacy.
Key Formulation Challenges for Clinical Translation
Translating biofilm-degrading enzymes from in vitro efficacy to in vivo therapeutics involves navigating a complex landscape of biological and physicochemical hurdles.
Enzyme Stability and Half-life: Proteins are susceptible to denaturation and proteolytic degradation in the harsh environment of infected tissues (e.g., the CF lung). Maintaining catalytic activity from production through storage and delivery to the site of infection is a primary challenge. Formulations must protect the enzyme's tertiary structure and active site integrity against pH extremes, host proteases, and reactive oxygen species.
Delivery and Penetration: The dense, anionic nature of the biofilm matrix itself impedes the diffusion of large molecules like enzymes. While enzymes degrade the matrix, their initial penetration into the biofilm's deep layers is limited. Furthermore, in the context of lung infection, effective delivery to the site of action requires overcoming the lung's clearance mechanisms. Formulation strategies such as encapsulation in nanoparticles or association with penetration-enhancing agents are being investigated to improve bioavailability and distribution.
Production and Scalability: Producing recombinant enzymes with high purity and consistent activity at a commercial scale requires optimized expression systems and rigorous quality control. The presence of even minor contaminants can trigger unwanted immune responses or exhibit off-target effects.
Immune Response and Safety: Repeated administration of bacterial-derived enzymes can be immunogenic. The potential for generating neutralizing antibodies that reduce therapeutic efficacy over time must be evaluated. Comprehensive toxicology studies are needed to ensure that enzyme activity does not damage host tissues, particularly given the structural similarities between some bacterial polysaccharides and human glycans.
Essential Experimental Protocols for Enzyme Characterization
Robust, standardized assays are critical for evaluating the efficacy of candidate enzymes and optimizing formulations. The following protocols are foundational to the field.
Biofilm Dissolution Assay with Microtiter Plates
Objective: To quantitatively measure the ability of an enzyme to dissolve pre-established biofilms. Materials: 96-well polystyrene plate, sterile growth medium (e.g., LB broth), crystal violet stain (0.2%), 96% ethanol, phosphate-buffered saline (PBS). Procedure:
- Biofilm Growth: Inoculate wells of the microtiter plate with 100 μL of bacterial suspension and incubate for 24-48 hours at 37°C to form a mature biofilm [59].
- Treatment: Carefully aspirate the planktonic cells and culture medium. Wash the biofilm gently with PBS to remove non-adherent cells. Add the enzyme solution in a suitable buffer to the test wells. Control wells receive buffer only.
- Incubation: Incubate the plate for a defined period (e.g., 2-24 hours) at 37°C.
- Staining and Quantification: Aspirate the enzyme solution, wash the wells, and stain the remaining adherent biofilm with 0.2% crystal violet for 15 minutes [59]. Wash excess stain away, solubilize the bound dye with 200 μL of 96% ethanol, and transfer 125 μL of the solution to a new plate.
- Data Analysis: Measure the absorbance of the solubilized crystal violet at 590 nm (A590) [59]. The percentage of biofilm dissolution is calculated by comparing the absorbance of enzyme-treated wells to the buffer-treated control wells.
Assessment of Antibiotic Synergy
Objective: To determine if a biofilm-degrading enzyme enhances the efficacy of a conventional antibiotic. Materials: Pre-established biofilms in a microtiter plate or flow cell, enzyme solution, antibiotic solution, viability stains (e.g., LIVE/DEAD BacLight kit) or materials for colony counting. Procedure:
- Treatment: Treat pre-established biofilms with: a) buffer control, b) enzyme alone, c) antibiotic alone, and d) enzyme and antibiotic in combination.
- Viability Assessment: After incubation, assess bacterial viability. This can be done by:
- Colony Forming Units (CFUs): Dislodge biofilm bacteria by sonication or scraping, serially dilute, and plate on agar to count CFUs after incubation.
- Confocal Microscopy: Use a flow cell system and stain with fluorescent viability dyes (e.g., SYTO9 and propidium iodide) to visualize live/dead cells in situ within the biofilm architecture [6].
- Data Analysis: Synergy is demonstrated when the combination treatment results in a statistically significant reduction in bacterial viability compared to either agent alone and the theoretical additive effect of both.
Figure 2: Experimental Workflow for Testing Enzyme-Antibiotic Synergy. This protocol tests the adjuvant potential of biofilm-degrading enzymes by comparing multiple treatment outcomes.
The Scientist's Toolkit: Research Reagent Solutions
Advancing research and development in this field relies on a suite of specialized reagents and tools. The following table catalogues essential materials for investigating biofilm-degrading enzymes.
Table 3: Key Research Reagent Solutions for Biofilm Enzyme Studies
| Reagent / Material | Function/Description | Example Application |
|---|---|---|
| Recombinant Alginate Lyases (e.g., Alg2A) | Enzymes that cleave alginate via β-elimination [58]. | Dissolving alginate-rich biofilms of mucoid P. aeruginosa; studying synergy with antibiotics [58]. |
| Psl Glycoside Hydrolase | Enzyme that specifically degrades the Psl polysaccharide. | Inhibiting initial attachment and disrupting Psl-dependent biofilm structures [20]. |
| Nanodiscs (e.g., MSP1D1) | Lipid bilayer discs for membrane protein reconstitution [9]. | Stabilizing and structurally analyzing membrane-embedded complexes like PelBC for Cryo-EM [9]. |
| Cryo-Electron Microscopy (Cryo-EM) | High-resolution structural determination technique for macromolecular complexes. | Solving the 3D structure of secretion complexes (e.g., PelBC at 2.5 Å) to understand export mechanisms [9]. |
| Particle Tracking Microrheology (PTM) | Technique to measure local viscoelastic properties of soft materials like biofilms. | Quantifying changes in biofilm stiffness and elasticity after enzyme treatment [6]. |
| Flow Cell Biofilm Reactor | System for growing biofilms under continuous medium flow for microscopic analysis. | Growing mature, structured biofilms for real-time, non-destructive analysis of enzyme effects [6]. |
The targeted degradation of the P. aeruginosa biofilm matrix using enzymes such as alginate lyases, Psl hydrolases, and prospective Pel-degrading enzymes represents a rational and promising strategy to overcome the pervasive challenge of biofilm-mediated antibiotic treatment failure. The path to clinical adoption hinges on overcoming significant drug delivery and formulation obstacles. Future success will rely on interdisciplinary collaboration between microbiology, structural biology, and pharmaceutical sciences to engineer stable, deliverable, and potent enzyme formulations. By integrating these novel adjuvants into the antimicrobial arsenal, the scientific community moves closer to effectively eradicating chronic P. aeruginosa infections and improving outcomes for vulnerable patient populations. The continued elucidation of EPS structures and their biosynthesis machineries, as demonstrated by recent Cryo-EM studies [9], will undoubtedly unveil new targets and inspire the next generation of anti-biofilm therapeutics.
Navigating Research and Therapeutic Challenges in EPS-Targeted Strategies
Pseudomonas aeruginosa thrives in diverse environments and poses significant challenges as an opportunistic pathogen, particularly in chronic infections such as those occurring in the cystic fibrosis (CF) lung [17] [16]. A key to its resilience is the biofilm mode of growth, wherein bacterial communities embed themselves in a self-produced extracellular matrix. This matrix provides structural integrity and confers formidable tolerance to antimicrobials and host defenses [16]. For P. aeruginosa, the extracellular polysaccharides (EPS)—alginate, Pel, and Psl—comprise a major component of this biofilm scaffold [17] [4]. While alginate is predominantly associated with mucoid variants isolated from chronic CF infections, non-mucoid strains utilize Pel, Psl, or a combination thereof to build their biofilms [17] [4].
A critical and often complicating feature in the biology of these biofilms is functional redundancy between the different EPS components. This redundancy presents a substantial obstacle for therapeutic strategies that aim to disrupt biofilms by targeting a single polysaccharide. This technical guide delves into the challenges inherent in targeting a single EPS within this multi-polysaccharide system, framing the discussion within the broader context of P. aeruginosa biofilm matrix research. We will synthesize current evidence, present structured experimental data, and outline methodologies to elucidate the mechanisms of redundancy and its implications for drug development.
The Roles and Regulation of Pel, Psl, and Alginate
The three exopolysaccharides—Pel, Psl, and alginate—are chemically distinct and play unique, yet sometimes overlapping, roles in biofilm formation and maintenance.
Psl is a neutral, branched pentasaccharide composed of D-mannose, D-glucose, and L-rhamnose repeats [17] [4]. It plays a critical role in the initial attachment of cells to surfaces and is involved in maintaining the structural integrity of the mature biofilm matrix [17] [16]. In the common lab strain PAO1, Psl serves as the primary structural scaffold, and its absence leads to a severe defect in surface attachment and biofilm development [17]. Psl is synthesized by proteins encoded by the 15-gene psl operon (pslA-O) [16].
Pel is a cationic polymer, rich in glucose and composed partially of N-acetylgalactosamine and galactosamine [16] [4]. Its name derives from its critical role in forming pellicles—biofilms at the air-liquid interface [4]. Pel facilitates cell-to-cell interactions and can cross-link with negatively charged extracellular DNA (eDNA) via ionic interactions, thereby contributing to biofilm cohesion [16]. In the strain PA14, which has a natural deletion in the psl operon, Pel is the primary structural polysaccharide [17]. The pel operon (pelA-G) is responsible for its synthesis [16].
Alginate is a high molecular weight, acidic polymer composed of O-acetylated D-mannuronic acid and L-guluronic acid [4]. It is the hallmark of mucoid P. aeruginosa phenotypes frequently isolated from CF lungs. While its overproduction contributes to the high viscosity of sputum in CF patients, it also forms a protective, gelatinous layer that shields the bacteria from host immune factors and antibiotics [16] [4].
Table 1: Characteristics of the Major P. aeruginosa Exopolysaccharides
| Polysaccharide | Chemical Composition | Primary Functions in Biofilm | Common Producing Strains |
|---|---|---|---|
| Psl | Neutral pentasaccharide (D-mannose, D-glucose, L-rhamnose) [17] [16] | Surface attachment, structural scaffold, cell-cell connections, antibiotic sequestration [17] [16] | Non-mucoid (e.g., PAO1) [17] |
| Pel | Cationic, partially de-N-acetylated (Galactosamine, N-acetylgalactosamine) [16] | Cell-cell interactions, pellicle formation, cross-links eDNA, scaffold in mature biofilms [17] [16] | Non-mucoid (e.g., PA14) [17] |
| Alginate | Acidic, non-repeating (D-mannuronic acid, L-guluronic acid), O-acetylated [4] | Matrix structural scaffold in mucoid strains, immune evasion, antibiotic tolerance [16] [4] | Mucoid CF isolates [4] |
The regulation of Pel and Psl is complex and intertwined. Key regulators include the transcriptional regulator FleQ, which represses both pel and psl operons. This repression is relieved by the secondary messenger cyclic di-GMP (c-di-GMP) [17]. Elevated intracellular c-di-GMP levels, often found in rugose small colony variants (RSCVs), lead to simultaneous upregulation of both Pel and Psl [17]. This coordinated regulation under a common signaling molecule is a foundational element of the functional redundancy observed in the system.
Figure 1: Simplified regulatory pathway for Pel and Psl expression. The secondary messenger c-di-GMP integrates environmental signals and relieves FleQ-mediated repression, leading to the transcription of the pel and psl operons and subsequent production of polysaccharides for the biofilm matrix.
Evidence for Functional Redundancy from Experimental Data
The concept of functional redundancy between Pel and Psl is not merely theoretical but is strongly supported by empirical evidence from studies of diverse P. aeruginosa isolates.
Strain-to-Strain Variability in EPS Dependence
A systematic analysis of clinical and environmental isolates revealed significant variability in the reliance on Pel or Psl for mature biofilm structure, leading to the proposal of four functional classes of strains [17]. While a psl mutation consistently impaired initial surface attachment across all strains tested, the impact of pel mutation on the mature biofilm structure was highly strain-dependent [17]. This suggests that while Psl's role in attachment is non-redundant, either Pel or Psl can fulfill the structural scaffold role in three-dimensional biofilm development, depending on the genetic background of the strain.
In Vivo Redundancy and Evolutionary Compensation
Perhaps the most compelling evidence for functional redundancy comes from evolution experiments. When a PAO1 psl mutant, which is initially deficient in biofilm formation, is subjected to extended cultivation, it can evolve to restore biofilm production [17]. This restoration occurs through mutations that lead to the constitutive upregulation of the otherwise silent pel operon [17]. This demonstrates that in the absence of the primary scaffold polysaccharide (Psl), the population can exploit the redundant capacity of the secondary polysaccharide (Pel) to regain a fitness advantage. This evolutionary compensation underscores the robustness that redundancy provides to the biofilm lifestyle.
Table 2: Phenotypic Consequences of pel and psl Mutations in Key P. aeruginosa Strains
| Strain / Genotype | Surface Attachment | Mature Biofilm Structure | Key Experimental Evidence |
|---|---|---|---|
| PAO1 (Wild-type) | Psl-dependent [17] | Psl-dependent [17] | psl mutation arrests development at monolayer stage [17] |
| PAO1 Δpel | Normal | Normal [17] | Biofilm biomass comparable to wild-type [17] |
| PAO1 Δpsl | Deficient | Severely impaired [17] | Biofilm arrested at monolayer; reduced biomass [17] |
| PAO1 ΔpelΔpsl | Deficient | Severely impaired [17] | Phenocopies Δpsl single mutant [17] |
| PA14 (Wild-type) | Psl-independent (has psl operon deletion) [17] | Pel-dependent [17] | pel mutation leads to reduced biofilm biomass and structure [17] |
| PA14 Δpel | Normal | Severely impaired [17] | Fails to form microcolonies in flow cells [17] |
| Evolved PAO1 Δpsl | Not Applicable | Restored | Acquires mutations that upregulate pel expression [17] |
Methodologies for Investigating EPS Redundancy
To study functional redundancy and its consequences, researchers employ a suite of well-established techniques. Below is a detailed protocol for a key experiment that directly demonstrates redundancy through evolutionary compensation.
Detailed Protocol: Experimental Evolution of Biofilm-Deficient Mutants
Objective: To determine whether a biofilm-deficient mutant (e.g., PAO1 Δpsl) can adapt during long-term culture to restore biofilm formation via upregulation of a redundant EPS pathway.
Materials and Reagents:
- Bacterial Strains: PAO1 wild-type, PAO1 Δpsl mutant, PAO1 Δpel mutant, PAO1 ΔpelΔpsl double mutant.
- Growth Medium: Dilute Lysogeny Broth (e.g., 1/10 or 1/20 LB) or other nutrient-limited media suitable for biofilm growth [17].
- Culture Vessels: Static microtiter plates (e.g., 96-well polystyrene plates) for initial attachment and biofilm biomass assays; flow-cell reactors for high-resolution structural analysis [17].
- Staining Reagents: Crystal violet solution (0.1%) for total biofilm biomass quantification [17].
- Molecular Biology Reagents: Primers for pel and psl operons, RNA extraction kit, reverse transcription kit, quantitative PCR (qPCR) reagents for gene expression analysis.
Procedure:
- Initial Phenotyping: Confirm the biofilm-deficient phenotype of the PAO1 Δpsl mutant using a microtiter dish biofilm assay [17].
- Grow strains overnight in liquid medium.
- Dilute cultures and inoculate into wells of a 96-well plate.
- Incubate statically for 8-24 hours at the desired temperature (e.g., 37°C).
- Remove planktonic cells, stain adherent biofilm with crystal violet, and destain with ethanol.
- Measure the optical density of the dissolved crystal violet to quantify relative biofilm biomass.
- Experimental Evolution Passaging:
- Inoculate the PAO1 Δpsl mutant into multiple independent flasks or wells containing a dilute biofilm-permissive medium.
- Incubate the cultures under static conditions for an extended period (e.g., 5-10 days) to allow for biofilm development and subsequent dispersal.
- Periodically (e.g., every 48-72 hours), passage the biofilm population by transferring a small aliquot of the culture (including both planktonic and biofilm cells) into fresh medium.
- Continue passaging for multiple cycles (e.g., 10-20 cycles) [17].
- Screening for Biofilm Restoration:
- At each passage, screen the evolved populations for restored biofilm formation using the microtiter dish assay described in Step 1.
- Islate clones from populations that show increased biofilm biomass compared to the ancestral Δpsl mutant.
- Confirmation of Redundancy Mechanism:
- Genetic Complementation: Attempt to complement the biofilm phenotype of the evolved isolate by reintroducing the wild-type psl operon. If complementation is unsuccessful, it suggests adaptation is via an alternative pathway.
- Gene Expression Analysis: Using qPCR, compare the transcription levels of the pel operon in the evolved isolate versus the ancestral Δpsl mutant. A significant upregulation of pel indicates a compensatory mechanism [17].
- Phenotypic Validation: Test the evolved isolate, and an evolved isolate with an introduced pel mutation, for biofilm formation. Loss of biofilm in the pel-deficient evolved isolate confirms that Pel is responsible for the restored phenotype [17].
- Structural Confirmation: Use confocal laser scanning microscopy (CLSM) of flow-cell-grown biofilms to visualize and compare the three-dimensional architecture of the wild-type, ancestral mutant, and evolved isolate biofilms [17].
Figure 2: Experimental workflow for evolving and characterizing EPS redundancy. The process involves serial passaging of a biofilm-deficient mutant, screening for restored biofilm formation, and validating the compensatory mechanism through molecular and microscopic techniques.
The Scientist's Toolkit: Essential Reagents for EPS Research
Table 3: Key Research Reagent Solutions for Investigating EPS Function and Redundancy
| Reagent / Tool | Function / Application | Example Use in Research |
|---|---|---|
| In-Frame Deletion Mutants (ΔpelF, ΔpslD) | Specifically abrogate production of Pel or Psl polysaccharide without polar effects, allowing for functional dissection [17]. | Used to determine the individual and combined contributions of Pel and Psl to attachment and biofilm structure in various strains [17]. |
| Quantitative PCR (qPCR) | Quantify transcript levels of pel and psl operon genes to measure regulation and expression under different conditions or in mutant backgrounds [17]. | Used to confirm upregulation of the pel operon in evolved PAO1 Δpsl isolates [17]. |
| Confocal Laser Scanning Microscopy (CLSM) | Generate high-resolution, three-dimensional images of hydrated live biofilms, allowing for visualization of structural phenotypes [17]. | Used to show that PA14 Δpel and PAO1 Δpsl mutants are arrested at the monolayer stage in flow-cells [17]. |
| Crystal Violet Biofilm Assay | High-throughput method to quantify total biofilm biomass attached to an abiotic surface in microtiter plates [17]. | Used for initial phenotyping of mutants and for screening evolved populations during experimental evolution [17]. |
| EPS-Specific Lectins / Antibodies | Fluorescently labeled probes that bind to specific EPS components, enabling spatial visualization within the biofilm matrix via microscopy [16]. | Used to stain and visualize the helical arrangement of Psl around PAO1 cells and its fabric-like matrix in biofilms [16] [4]. |
| Flow-Cell Reactor System | Cultivate biofilms under continuous nutrient flow, which promotes development of complex 3D structures reminiscent of natural environments [17]. | The gold-standard system for analyzing the architecture of mature biofilms formed by wild-type and EPS mutant strains [17]. |
Challenges and Considerations for Therapeutic Targeting
The demonstrated functional redundancy between Pel and Psl has profound implications for developing anti-biofilm therapies that target EPS components.
The Risk of Compensatory Evolution: Targeting a single EPS, such as Psl with a specific glycoside hydrolase enzyme, may successfully disrupt the biofilm of a strain that primarily relies on that polymer. However, this intervention imposes a strong selective pressure that can favor pre-existing or de novo mutants that upregulate the redundant Pel polysaccharide [17]. This compensatory evolution would lead to therapeutic failure and potentially select for more resilient biofilm-forming variants.
Strain Heterogeneity in Clinical Settings: Clinical infections, particularly chronic CF lung infections, are often composed of a heterogeneous population of P. aeruginosa strains and variants [16]. This population can include a mix of mucoid (alginate-producing) and non-mucoid isolates, as well as RSCVs that hyper-produce Pel and/or Psl [17] [16]. A mono-therapy targeting one EPS is unlikely to be effective against such a diverse population, as different subpopulations rely on different EPS for biofilm integrity.
The Redundancy in Protective Functions: Beyond their structural roles, EPS components also contribute redundantly to antibiotic tolerance. Both Psl and Pel have been shown to contribute to tolerance against aminoglycoside antibiotics like tobramycin, potentially through cation sequestration mechanisms [16]. Disabling one EPS may not sufficiently increase antibiotic susceptibility if another redundant EPS can perform a similar protective function.
The functional redundancy between Pel, Psl, and alginate within the P. aeruginosa biofilm matrix represents a sophisticated survival strategy. It ensures robustness and resilience, allowing the bacterial community to maintain its protective biofilm structure in the face of genetic mutations or environmental perturbations, including targeted anti-biofilm agents. The evolutionary experiment where a psl mutant resuscitates biofilm formation by activating Pel production is a powerful illustration of this concept [17].
Moving forward, overcoming the challenge of functional redundancy requires a paradigm shift in therapeutic strategy. Future research and drug development efforts should move beyond single-target approaches and consider:
- Multi-Targeted Therapies: Developing combination therapies that simultaneously target two or more EPS systems (e.g., a Psl-degrading enzyme combined with a Pel-disrupting agent) could prevent compensatory upregulation and lead to more effective biofilm dispersal.
- Targeting Central Regulators: Interventions aimed at the core regulatory pathways common to all EPS systems, such as the c-di-GMP signaling network, hold promise for broadly suppressing matrix production without triggering redundant pathways [17].
- Personalized Medicine Approaches: Diagnosing the specific EPS expression profile of a patient's infecting P. aeruginosa population could guide the selection of the most appropriate targeted therapy, much like antibiotic susceptibility testing guides antibiotic choice.
Understanding the intricacies of functional redundancy is not merely an academic exercise; it is a critical step toward designing the next generation of effective anti-biofilm treatments for one of the most persistent and problematic pathogens in modern medicine.
The extracellular polymeric matrix of Pseudomonas aeruginosa biofilms, primarily composed of the exopolysaccharides alginate, Psl, and Pel, constitutes a primary determinant of virulence and antibiotic resistance in chronic infections such as those in cystic fibrosis (CF) lungs. However, the accurate detection, spatial discrimination, and quantification of these matrix components in situ present significant diagnostic challenges that impede the development of targeted therapies. This whitepaper delineates the specific structural and functional roles of alginate, Psl, and Pel within the biofilm matrix, highlighting the critical limitations of current analytical methods. We further synthesize experimental data on the phenotypic consequences of disrupting each polysaccharide and propose a framework for the development of next-generation specific molecular probes and non-invasive in vivo detection technologies. Overcoming these diagnostic hurdles is essential for advancing our fundamental understanding of biofilm architecture and accelerating the clinical translation of matrix-targeting anti-biofilm strategies.
Pseudomonas aeruginosa biofilm infections are notoriously difficult to eradicate, largely due to the protective extracellular matrix that encases the bacterial communities [2]. This matrix provides structural integrity, mediates adhesion, and imposes a formidable barrier to antibiotics and host immune defenses [6] [60]. The three self-secreted exopolysaccharides—alginate, Psl, and Pel—are core components of this matrix, and their expression varies significantly between strains and in response to environmental cues [17] [4].
The transition to chronic infection in the CF lung is often marked by the emergence of mucoid P. aeruginosa variants that overproduce alginate, a polymer of D-mannuronic and L-guluronic acids [2] [60]. In contrast, non-mucoid strains, including common laboratory models like PAO1 and PA14, typically utilize Psl, a neutral pentasaccharide of D-mannose, D-glucose, and L-rhamnose, and/or Pel, a cationic polymer of partially de-N-acetylated N-acetylgalactosamine, as their primary structural scaffolds [17] [2] [4]. The ability to specifically identify and localize these EPS components within a complex, host-derived milieu is a prerequisite for disassembling the biofilm matrix and restoring antibiotic efficacy. Current methods, which largely rely on in vitro models, fail to recapitulate the chemical complexity and heterogeneity of in vivo biofilms, creating a critical diagnostic gap [61].
Distinct Roles and Diagnostic Challenges of Core Exopolysaccharides
The functional redundancy and unique properties of alginate, Psl, and Pel necessitate detection methods capable of high specificity. The following sections detail their distinct roles and the associated diagnostic challenges.
Alginate: The Mucoid Shield
- Structure and Function: Alginate is a high molecular weight, anionic, and often O-acetylated linear copolymer of β-D-mannuronic acid and its C5 epimer α-L-guluronic acid [4] [60]. In mucoid strains, it forms a dense, viscous, gelatinous capsule that surrounds cells, contributing significantly to the mechanical stability of biofilms and providing a potent diffusion barrier against antibiotics and reactive oxygen species from neutrophils [2] [6].
- Diagnostic Challenge: The overproduction of alginate in mucoid variants is a key clinical indicator. However, its physical density and chemical similarity to host mucins complicate specific targeting and imaging within sputum or lung tissue. Probes must distinguish bacterial alginate from human polyanionic glycans.
Psl: The Structural Architect
- Structure and Function: Psl is a neutral, branched pentasaccharide composed of repeating units of D-mannose, L-rhamnose, and D-glucose [17] [2]. It is critical for surface attachment, cell-cell interactions, and the formation of the biofilm's structural core. In strain PAO1, Psl acts as the primary scaffold, creating a "fabric-like" matrix that connects cells [17] [4]. It also plays a key role in biofilm elasticity and thickness [8].
- Diagnostic Challenge: Psl's helical arrangement on the cell surface and its role in initial attachment make it an ideal target for early-diagnostic and preventive strategies. However, its neutral charge and structural heterogeneity require probes that can recognize its unique pentasaccharide repeating unit without cross-reacting with other polysaccharides.
Pel: The Protective Cationic Polymer
- Structure and Function: Pel is a cationic linear polymer of α-1-4-linked N-acetyl-D-galactosamine that is partially deacetylated during its secretion, conferring a positive charge [2] [9]. This charge allows it to cross-link with negatively charged extracellular DNA (eDNA) within the matrix, strengthening the biofilm structure [2]. Pel is crucial for pellicle formation and, in strains like PA14, serves as the primary structural polysaccharide [17] [3]. A key protective function of Pel is the sequestration of positively charged aminoglycoside antibiotics, such as tobramycin, rendering the biofilm more tolerant [3].
- Diagnostic Challenge: The cationic nature of Pel is unique among the major EPS. Diagnostics that exploit this charge for detection must be designed to function accurately in the complex ionic environment of an infection site, where other polycations may be present.
Table 1: Core Exopolysaccharides of P. aeruginosa Biofilms
| Exopolysaccharide | Chemical Composition | Charge | Primary Functions | Strain Example |
|---|---|---|---|---|
| Alginate | D-mannuronic acid, L-guluronic acid [60] | Anionic [6] | Mechanical stability, antibiotic barrier, immune evasion [2] [60] | Mucoid CF isolates (e.g., FRD1) [4] |
| Psl | D-mannose, L-rhamnose, D-glucose [17] [2] | Neutral | Surface attachment, primary structural scaffold, biofilm elasticity [17] [8] | PAO1 [17] [8] |
| Pel | Partially de-N-acetylated N-acetylgalactosamine [2] [9] | Cationic [2] | Cell-cell interactions, aminoglycoside sequestration, structural redundancy [17] [3] | PA14 [17] [3] |
Quantitative Phenotypic Analysis of EPS-Deficient Mutants
Systematic mutational analysis reveals the distinct and overlapping contributions of each EPS to the biofilm phenotype. Recent multi-omics studies provide quantitative data on the phenotypic consequences of disrupting each biosynthesis pathway.
Table 2: Phenotypic Characterization of P. aeruginosa PAO1 EPS Mutants
| Biofilm Parameter | PAO1-WT | PAO1-ΔpelF | PAO1-ΔpslAB | PAO1-Δalg8 |
|---|---|---|---|---|
| Biofilm Biomass (Crystal Violet) | 100% | ~33% [8] | <30% [8] | No significant change [8] |
| Biofilm Thickness | Baseline | Reduced [8] | ~50% reduction [8] | No significant change [8] |
| Elastic Modulus | Baseline | Slight reduction | Significant reduction [8] | No significant change [8] |
| Primary Functional Loss | - | Redundant structural support [17] | Structural integrity & attachment [17] [8] | Alginate production [8] |
Data derived from multi-omics analysis of PAO1 mutants, showing the hierarchical importance of Psl for biofilm formation in this strain [8]. The deletion of pslAB has the most dramatic effect, while the loss of alg8 has a minimal impact, consistent with the non-mucoid nature of standard PAO1.
The data in Table 2 underscores a critical concept in biofilm biology: functional redundancy. While Psl is the dominant scaffold in PAO1, the Pel polysaccharide can compensate for its absence. This was elegantly demonstrated in an evolution experiment where a PAO1 psl mutant, initially producing a poor biofilm, acquired compensatory mutations that upregulated pel expression after extended cultivation, thereby restoring biofilm-forming capacity [17]. This redundancy is a major survival advantage for the bacterium and a significant complication for monotherapeutic strategies that target only a single EPS.
Experimental Approaches for EPS Disruption and Analysis
Protocol: Assessing Biofilm Phenotypes Using Microtiter Dish Assays and Confocal Microscopy
This fundamental protocol is used to quantify attachment and biofilm formation deficits in EPS mutants [17] [3].
- Strain Preparation: Create in-frame, non-polar deletion mutants of key biosynthetic genes (e.g.,
pelF,pslD,pslAB) in the desired genetic background (e.g., PAO1, PA14) using allelic replacement [17]. Include complementary overexpression strains. - Growth Normalization: Grow all test strains in appropriate liquid medium (e.g., LB) to mid-log phase. Normalize the cell density optically (e.g., OD600) to ensure equivalent inoculation levels [17].
- Microtiter Dish Assay (Attachment):
- Transfer normalized cell suspensions to a 96-well polystyrene microtiter plate.
- Incubate statically for a short period (e.g., 2-4 hours) at the desired temperature.
- Gently wash wells with buffer to remove non-adherent cells.
- Fix attached cells with methanol or ethanol and stain with a 0.1% crystal violet solution for 15 minutes.
- Wash away excess stain, solubilize the bound crystal violet in acetic acid or ethanol, and measure the absorbance at 590 nm to quantify attachment [17] [3].
- Microtiter Dish Assay (Mature Biofilm): Repeat step 3, but extend the incubation time to 24-48 hours to allow for mature biofilm development before washing and staining [3].
- Confocal Laser Scanning Microscopy (CLSM) Analysis:
- Grow biofilms in flow cells or on coverslips to preserve the 3D structure.
- Use strains constitutively expressing a fluorescent protein (e.g., GFP) for cell visualization.
- For EPS-specific staining, apply fluorescently labelled lectins or antibodies (e.g., Psl-specific lectin) or use DNA-binding dyes (e.g., TOTO-1) to label eDNA that interacts with Pel.
- Acquire z-stack images of the biofilm using a CLSM.
- Use image analysis software (e.g., COMSTAT, ImageJ) to quantify biovolume, thickness, and roughness coefficient [17] [60].
Protocol: Evaluating Anti-Biofilm Efficacy of EPS-Targeting Agents
This protocol assesses the ability of compounds like alginate oligosaccharides (OligoG) to disrupt established biofilms [60].
- Biofilm Establishment: Grow a mucoid P. aeruginosa biofilm (e.g., strain NH57388A) in a flow cell or on a coverslip for 24-72 hours to form a mature, structured biofilm.
- Treatment with Agent:
- Prepare solutions of the anti-biofilm agent (e.g., OligoG CF-5/20 at 0.5%-6% w/v) in an appropriate buffer or medium.
- For established biofilm disruption, carefully apply the treatment solution to the pre-formed biofilm and incubate for a set period (e.g., 1-24 hours) [60].
- Include an untreated control biofilm incubated with buffer alone.
- Viability and Biomass Assessment:
- Use live/dead bacterial viability staining (e.g., SYTO 9/propidium iodide) in conjunction with CLSM to assess cell viability within the biofilm post-treatment.
- Quantify the reduction in biofilm biomass and thickness using COMSTAT analysis of CLSM z-stacks [60].
- Potentiation of Antibiotics:
- Co-administer a sub-inhibitory concentration of the anti-biofilm agent with a conventional antibiotic (e.g., tobramycin, colistin).
- Compare the reduction in viable biofilm counts (CFU) to the effects of the antibiotic alone to demonstrate synergy [60].
Visualization of EPS Biosynthesis and Detection Challenges
The following diagram illustrates the biosynthesis and functional relationships between the three exopolysaccharides, highlighting the critical junctures where specific detection remains a challenge.
Diagram Title: EPS Biosynthesis and Detection Challenges
This diagram maps the intracellular biosynthesis and export pathways of alginate, Psl, and Pel to their final functional roles in the extracellular matrix. The distinct chemical properties of each polymer—alginate (anionic, gel-forming), Psl (neutral, structural scaffold), and Pel (cationic, cross-linking)—are key to their function but also complicate specific detection. A central diagnostic challenge, highlighted in red, is the current lack of molecular probes that can reliably distinguish between these EPS components within the complex, host-derived environment of an in vivo infection site.
The Scientist's Toolkit: Key Research Reagents and Models
Advancing diagnostics and therapeutics for P. aeruginosa biofilms relies on a specific toolkit of reagents, mutant strains, and model systems.
Table 3: Essential Research Tools for EPS-Focused Biofilm Research
| Tool / Reagent | Function / Description | Key Application |
|---|---|---|
| Defined Mutant Strains | Isogenic mutants with deletions in key genes (e.g., pslAB, pelF, alg8) [17] [8]. |
Phenotypic comparison to determine the specific role of each EPS in biofilm formation and architecture. |
| EPS-Specific Lectins/Antibodies | Fluorescently tagged molecular probes that bind to specific EPS components (e.g., Psl-specific lectin) [4]. | Visualization and spatial localization of specific polysaccharides within the biofilm matrix using CLSM. |
| Alginate Oligosaccharides (OligoG) | Low molecular weight alginate derivatives (Mn ~3200 g/mol) that disrupt the EPS matrix [60]. | Studying matrix disruption, antibiotic potentiation, and diffusion dynamics within biofilms. |
| Ex Vivo Pig Lung (EVPL) Model | A clinically realistic biofilm platform using porcine bronchiolar tissue [61]. | Bridge the gap between in vitro studies and CF lung infection; validates pathways (e.g., GacA, PelA) essential for in vivo-like biofilm formation [61]. |
| Particle Tracking Microrheology (PTM) | A non-invasive technique using embedded microparticles to measure local viscoelastic properties [6]. | Quantifying the mechanical contributions of different EPS components (e.g., alginate increases elastic modulus) to biofilm stability [6]. |
The hierarchical and often redundant functions of alginate, Psl, and Pel in P. aeruginosa biofilms represent a sophisticated bacterial defense strategy. The quantitative data clearly show that while Psl can be the dominant structural scaffold in strains like PAO1, Pel provides a crucial backup system, and alginate defines the resistance of mucoid variants [17] [8]. This functional interplay, combined with the starkly different chemical nature of each polymer, creates a complex diagnostic problem. Current in vitro models and detection methods are insufficient to resolve the spatial and temporal dynamics of these components in real infections.
The path forward requires a concerted effort in chemical biology and biomedical engineering to develop a new generation of diagnostics. Priority areas include the synthesis of high-affinity, optically active probes (e.g., lectins, antibodies, or aptamers) that can distinguish Psl from Pel and alginate within a host environment. Furthermore, these probes must be compatible with non-invasive imaging modalities suitable for clinical translation. Success in this endeavor will not only refine our basic understanding of biofilm pathology but will also provide the critical tools needed to monitor the efficacy of matrix-targeting therapies in real-time, ultimately paving the way for more effective treatments for chronic P. aeruginosa infections.
The biofilm matrix of Pseudomonas aeruginosa constitutes a formidable barrier in the treatment of chronic infections, particularly in cystic fibrosis (CF) lungs. This self-secreted milieu is a complex amalgamation of extracellular polymeric substances, with the exopolysaccharides (EPS) alginate, Psl, and Pel serving as foundational architectural components [2]. These EPS provide structural integrity to biofilms and confer significant tolerance to antimicrobials and host immune defenses [2] [3]. Current therapeutic strategies increasingly focus on enzymatic disruption of this matrix as an adjuvant to conventional antibiotics. However, the efficacy of enzyme-based treatments is governed by their stability, specificity, and penetration capacity within the intricate biofilm environment. This technical guide examines the core properties of P. aeruginosa EPS and outlines advanced methodologies for developing and evaluating enzymes capable of navigating the complex biofilm milieu to potentiate conventional antimicrobial therapies.
Structural and Functional Landscape of P. aeruginosa Exopolysaccharides
Chemical Composition and Biosynthesis
P. aeruginosa produces three primary exopolysaccharides—alginate, Psl, and Pel—each with distinct chemical structures and biosynthetic pathways that dictate their functional roles within the biofilm matrix.
Alginate is a high molecular weight, acidic polysaccharide composed of non-repeating subunits of D-mannuronic acid and its C5' epimer L-guluronic acid, often O-acetylated which enhances its protective properties [4]. It is typically overproduced by mucoid variants isolated from chronically infected CF lungs, where it forms a viscous, gelatinous layer surrounding bacterial cells [2] [4].
Psl is a neutral, branched pentasaccharide composed of repeating D-mannose, D-glucose, and L-rhamnose subunits [2] [4]. Its synthesis involves 15 co-transcribed genes (pslA-O), and it forms a helical arrangement on the cell surface that facilitates surface attachment and cell-cell interactions [2]. Visualization through confocal laser scanning microscopy reveals Psl forms a fabric-like matrix connecting biofilm cells [4].
Pel is a cationic polymer composed of partially deacetylated α-1,4-linked N-acetylgalactosamine (GalNAc) residues [9] [12]. This positive charge enables ionic cross-linking with negatively charged extracellular DNA (eDNA) within the biofilm matrix [2]. The pel operon (pelA-G) encodes its biosynthesis machinery, with recent structural studies revealing detailed mechanisms of its export through the outer membrane PelBC complex [9].
Table 1: Comparative Analysis of P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Charge Properties | Primary Biosynthetic Genes | Structural Role in Biofilm |
|---|---|---|---|---|
| Alginate | D-mannuronic acid, L-guluronic acid | Acidic | alg genes | Forms viscous capsule; dominant in mucoid CF isolates |
| Psl | D-mannose, D-glucose, L-rhamnose (pentasaccharide repeat) | Neutral | pslA-O (15 genes) | Cell-surface helices; primary scaffold in PAO1 strains |
| Pel | Partially de-N-acetylated GalNAc (α-1,4-linked) | Cationic | pelA-G (7 genes) | Pellicle formation; primary scaffold in PA14 strains |
Strain-Specific Expression and Functional Redundancy
The relative contribution of each exopolysaccharide to biofilm formation exhibits significant strain-dependent variability. Laboratory strain PAO1 relies primarily on Psl as its structural scaffold, while strain PA14 (which has a natural deletion in parts of the psl operon) depends predominantly on Pel [17] [3]. This functional redundancy was demonstrated through evolution experiments where PAO1 psl mutant strains, after extended cultivation, acquired compensatory mutations that upregulated Pel expression, restoring biofilm formation capacity [17].
The expression of these EPS is regulated by complex networks including transcriptional control by FleQ (repressed by c-di-GMP), RpoS, quorum sensing systems, and the Gac-Rsm signal transduction pathway [17]. Rugose small colony variants (RSCVs), frequently isolated from CF sputum, exhibit hyperadherence and hyperaggregation phenotypes resulting from elevated c-di-GMP signaling and consequent simultaneous overexpression of both Pel and Psl [17].
Protective Functions in Biofilm Environments
Beyond their structural roles, these exopolysaccharides confer critical protective functions:
- Antibiotic sequestration: Pel confers specific tolerance to aminoglycoside antibiotics such as tobramycin through electrostatic interactions with its cationic polymers, while Psl provides broader-spectrum antibiotic exclusion [2] [3].
- Immune evasion: Psl prevents efficient complement deposition and inhibits neutrophil reactive oxygen species production and phagocytosis [2].
- Matrix stabilization: Pel-eDNA interactions protect eDNA from degradation by DNase I, potentially rendering nuclease-based therapies less effective [2].
Enzymatic Targeting of Biofilm Exopolysaccharides: Current Approaches
Enzyme Classes and Mechanisms
The distinct chemical properties of each exopolysaccharide necessitate tailored enzymatic approaches for matrix disruption.
Alginate lyases cleave the glycosidic bonds between mannuronic and guluronic acid residues through a β-elimination mechanism, effectively depolymerizing the alginate matrix and reducing biofilm integrity [20]. These enzymes are particularly relevant for targeting mucoid P. aeruginosa isolates prevalent in CF infections.
Psl glycoside hydrolases target the pentasaccharide repeating units of Psl, disrupting its role in initial attachment and biofilm structural stability [20]. The neutral charge of Psl requires hydrolases with specific affinity for its mannose-glucose-rhamnose composition.
Pel-deacetylating enzymes and Pel hydrolases represent emerging approaches targeting the Pel matrix. PelA demonstrates dual functionality with both deacetylase and hydrolase (endo-α-1,4-N-acetylgalactosaminidase) activities [12]. The deacetylation process is crucial for generating the positive charge that facilitates Pel's interaction with eDNA, while the hydrolase activity directly cleaves the polysaccharide backbone.
Table 2: Enzyme-Based Strategies Against P. aeruginosa Exopolysaccharides
| Target EPS | Enzyme Class | Catalytic Mechanism | Key Challenges | Experimental Evidence |
|---|---|---|---|---|
| Alginate | Alginate lyase | β-elimination of glycosidic bonds | Effective mainly against mucoid strains | Reduces biofilm viscosity; enhances antibiotic penetration |
| Psl | Psl glycoside hydrolase | Hydrolysis of pentasaccharide repeats | Limited efficacy against Pel-dominated biofilms | Inhibits biofilm formation on lung epithelial cells [20] |
| Pel | PelA (deacetylase/hydrolase) | De-N-acetylation & glycosidic bond cleavage | Cationic nature may hinder enzyme access | Structural analysis at 2.1Å; enhanced antibiotic susceptibility [12] |
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for Biofilm EPS Studies
| Reagent/Resource | Function/Application | Experimental Utility |
|---|---|---|
| Isogenic mutant strains (e.g., Δpel, Δpsl, ΔpelΔpsl) | Genetic dissection of individual EPS functions | Controlled studies of specific EPS contributions to biofilm architecture and resistance |
| PDO300 (MucA-negative mutant) | Constitutive alginate overproduction | Models mucoid conversion in CF infections |
| PAO1PBADpel / PA14PBADpel | Arabinose-inducible Pel expression | Tunable systems for studying Pel dosage effects |
| Congo red binding assay | Polysaccharide detection and quantification | Rapid assessment of EPS production levels |
| PelBC complex in nanodiscs | Membrane protein structural studies | Cryo-EM analysis of Pel export machinery [9] |
| Flow-cell reactor with confocal microscopy | Real-time biofilm visualization | Spatial analysis of biofilm architecture and enzyme penetration |
Methodological Framework for Evaluating Enzyme Efficacy
Experimental Workflows for Enzyme Characterization
The following diagram illustrates a comprehensive workflow for evaluating anti-biofilm enzyme efficacy, from biochemical characterization to functional assessment:
Detailed Protocols for Key Assays
Microtiter Dish Biofilm Assay
This foundational assay quantifies biofilm formation on plastic surfaces during static incubation [17] [3]:
- Inoculum preparation: Grow P. aeruginosa overnight in appropriate medium (e.g., LB), dilute to ~1×10^6 CFU/mL in fresh medium.
- Enzyme treatment: Add purified enzyme at varying concentrations (0.1-100 μg/mL) to wells containing bacterial inoculum.
- Incubation: Static incubation at desired temperature (typically 37°C) for 24-48 hours.
- Staining and quantification: Remove planktonic cells, fix biofilms with methanol, stain with 0.1% crystal violet for 15 minutes, wash, solubilize dye in acetic acid/ethanol, measure absorbance at 595 nm.
- Controls: Include untreated controls, enzyme-only controls (to assess effect on pre-formed biofilms), and antibiotic combination treatments.
Flow-Cell Reactor with Confocal Laser Scanning Microscopy (CLSM)
This system enables real-time visualization of biofilm structural dynamics under conditions mimicking natural environments [17] [3]:
- System setup: Assemble flow-cell chambers with appropriate substratum (glass, polycarbonate, or medically-relevant materials).
- Conditioning and inoculation: Condition with medium for 1 hour, inoculate with bacterial suspension (1×10^7 CFU/mL in medium), allow attachment without flow for 1-2 hours.
- Continuous flow: Initiate continuous medium flow (typically 0.1-0.2 mm/s) to establish nutrient supply and shear forces.
- Enzyme treatment: Introduce enzyme treatments after specified biofilm development time (e.g., 24-72 hours) via injection or continuous infusion.
- Staining and imaging: Stain with appropriate fluorescent markers (e.g., SYTO9 for cells, lectins for specific polysaccharides, propidium iodide for viability), image using CLSM with appropriate laser settings and filter sets.
- Image analysis: Use software such as IMARIS, COMSTAT, or ImageJ to quantify biomass, thickness, roughness coefficient, and substratum coverage.
Reverse Osmosis Membrane Biofouling System
Specialized system for evaluating biofouling and dispersal under hydraulic flow conditions [18]:
- Membrane conditioning: Condition polyamide RO membranes with artificial wastewater or appropriate medium.
- Biofilm development: Circulate bacterial inoculum through cross-flow membrane cells under pressure (typically 50-100 psi).
- Performance monitoring: Track transmembrane pressure (TMP) and permeate flux to assess biofouling development.
- Enzyme treatment: Introduce enzymes at various biofilm maturation stages.
- Dispersal assessment: Quantify dispersed cells in permeate and retentate streams, analyze membrane surfaces using microscopy and EPS extraction.
Advanced Optimization Strategies
Engineering Enhanced Enzyme Stability
The biofilm microenvironment presents numerous challenges to enzyme stability, including protease activity, oxidative stress, and pH fluctuations. Strategic approaches to enhance stability include:
- Glycosylation engineering: Modifying surface glycosylation patterns to protect against proteolytic degradation.
- Cationic charge modulation: For enzymes targeting anionic EPS like alginate, optimizing surface charge to enhance substrate interaction while maintaining solubility.
- Thermostability engineering: Introducing strategic mutations to increase melting temperature (Tm) for enhanced longevity under infection conditions.
- Domain fusion strategies: Creating chimeric enzymes with multiple catalytic activities (e.g., combining alginate lyase with Psl hydrolase activities) for broader matrix disruption.
Pathfinding and Penetration Enhancement
The dense, heterogeneous structure of biofilms significantly impedes enzyme penetration. Advanced strategies to overcome this limitation include:
- Size optimization: Engineering smaller, catalytically active enzyme fragments (<30 kDa) to enhance diffusion through matrix pores.
- Synergistic cocktails: Combining EPS-degrading enzymes with DNases (e.g., recombinant human DNase I) to disrupt complementary matrix components [2].
- Delivery system engineering: Utilizing nanoparticle-based carriers that release enzymes in response to biofilm-specific stimuli (low oxygen, high cell density).
- Quorum sensing disruption: Combining matrix degradation with quorum quenching agents to simultaneously inhibit biofilm development and disperse established communities.
The strategic disruption of P. aeruginosa biofilm matrices through enzymatic targeting of alginate, Psl, and Pel represents a promising adjuvant approach to conventional antibiotic therapies. The distinct chemical properties, biosynthetic pathways, and functional roles of each exopolysaccharide necessitate tailored enzymatic strategies with optimized stability, specificity, and penetration capabilities. Comprehensive evaluation using the methodological framework outlined in this guide—spanning biochemical characterization, in vitro biofilm models, and advanced efficacy assessment—enables rational development of next-generation enzyme-based therapeutics. As research advances, particularly in structural biology revealing the molecular machinery of EPS assembly and export [9] [12], opportunities emerge for designing increasingly sophisticated interventions to overcome the formidable barrier presented by the complex biofilm milieu.
The extracellular polymeric substance (EPS) matrix is a defining feature of bacterial biofilms, serving as both a structural scaffold and a protective barrier. In the opportunistic pathogen Pseudomonas aeruginosa, this matrix can consist of three primary exopolysaccharides: alginate, Psl, and Pel. A key survival strategy employed by P. aeruginosa is the compensatory upregulation of alternative EPS components when the production of one is disrupted. This adaptive resistance mechanism ensures the structural integrity and defensive capabilities of the biofilm are maintained, posing a significant challenge in clinical settings, particularly in the treatment of chronic infections such as those in cystic fibrosis (CF) airways. This whitepaper delves into the molecular basis of this compensation, synthesizing current research to provide a technical guide for scientists and drug development professionals aiming to disrupt these resilient biofilm structures.
Structural and Functional Roles of Core Exopolysaccharides
P. aeruginosa produces three main exopolysaccharides—alginate, Psl, and Pel—each with a distinct chemical structure and functional role within the biofilm matrix. A comparative overview of their characteristics is provided in the table below.
Table 1: Characteristics of Core P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Primary Genetic Locus | Key Functions in Biofilm | Strain Association |
|---|---|---|---|---|
| Alginate | Acetylated copolymer of D-mannuronic and L-guluronic acids [1] [16] | alg genes (e.g., alg8) [14] | Protection from ROS and phagocytosis; confers resistance in chronic CF infections [1] [62] | Mucoid variants (e.g., CF isolates) [1] |
| Psl | Repeating pentasaccharide of D-mannose, D-glucose, and L-rhamnose [16] [4] | psl operon (pslA-O) [1] [16] | Primary scaffold in non-mucoid strains; critical for surface attachment, cell-cell interactions, and biofilm elasticity [1] [14] [17] | Non-mucoid strains (e.g., PAO1) [17] |
| Pel | Cationic, partially de-N-acetylated polymer of N-acetylgalactosamine and N-acetylglucosamine [63] [16] | pel operon (pelA-G) [1] [16] | Cell-cell adhesion, pellicle formation, and tobramycin sequestration via eDNA cross-linking [1] [16] | Non-mucoid strains (e.g., PA14) [17] |
Experimental Evidence for EPS Compensation
Phenotypic and Fitness Outcomes of EPS Deletion
Multi-omics approaches and targeted mutational analyses have delineated the hierarchical roles and compensatory potential of the EPS components. Recent multi-omics research demonstrates that deletion of the primary structural polysaccharide leads to significant phenotypic shifts and metabolic reprogramming, while the absence of non-dominant EPS has minimal impact [14]. Key quantitative findings from phenotypic characterization are summarized below.
Table 2: Phenotypic Impact of EPS Gene Deletions in P. aeruginosa PAO1
| Parameter | PAO1-ΔpslAB | PAO1-ΔpelF | PAO1-Δalg8 |
|---|---|---|---|
| Biofilm Biomass (vs. WT) | Decreased to <⅓ of WT [14] | Reduced to ~⅓ of WT [14] | No significant difference [14] |
| Biofilm Architecture | Significant reduction; dispersed cells [14] | Notable reduction [14] | Similar to WT; lacks external filaments [14] |
| Elastic Modulus | ~4x increase (more rigid) [14] | Not significantly different [14] | Not significantly different [14] |
| Role in PAO1 | Essential for structure and attachment [14] [17] | Redundant, reinforces structure [14] | Minimal impact on formation [14] |
Despite these clear phenotypic deficits in vitro, the loss of Pel and/or Psl does not always alter the overall bacterial load or rate of wound closure in a murine infection model [63]. However, the spatial distribution of bacterial aggregates within the wound tissue is significantly affected, with ΔpelΔpsl double mutants forming much smaller aggregates spaced farther apart [63]. Crucially, the loss of both Pel and Psl significantly impairs the ability of P. aeruginosa to survive aminoglycoside treatment in vivo, underscoring the critical role of these EPS in antimicrobial tolerance [63].
Molecular Mechanisms of Adaptive EPS Upregulation
The compensatory upregulation of Pel in a PAO1 psl mutant is a canonical example of adaptive resistance at the genetic level. In this common laboratory strain, which primarily utilizes Psl, the deletion of the psl operon initially results in a biofilm-deficient phenotype. However, after extended cultivation, these psl mutant populations acquire mutations that lead to the upregulation of the Pel polysaccharide [17]. This functional redundancy between Pel and Psl as structural scaffolds helps preserve biofilm formation capacity when primary EPS genes are compromised.
This compensation is often mediated through mutations in regulatory pathways that control the intracellular levels of the secondary messenger cyclic di-GMP (c-di-GMP). For instance, loss-of-function mutations in the wspF gene are a common genetic route to this compensatory phenotype [1] [17]. WspF is a negative regulator of the diguanylate cyclase WspR. Therefore, wspF mutations lead to constitutive activation of WspR, resulting in elevated global c-di-GMP levels [1] [17]. High c-di-GMP, in turn, promotes the transcription of the pel operon, among other biofilm-promoting genes, thereby compensating for the absence of Psl [17].
Diagram: Regulatory Pathway for EPS Compensation via Wsp System
Methodologies for Investigating EPS Compensation
Essential Experimental Workflow
A combination of genetic, phenotypic, and biochemical assays is required to conclusively demonstrate EPS compensation. The following workflow outlines the key stages of a robust experimental design.
Diagram: Experimental Workflow for Characterizing EPS Compensation
Detailed Methodologies for Key Assays
4.2.1 Genetic Mutagenesis and Strain Construction
- Targeted Gene Deletion: Generate in-frame, markerless deletions of key EPS biosynthesis genes (e.g., pslD for Psl, pelF for Pel, alg8 for alginate) using allelic exchange with a suicide vector, such as pEX18Gm [14] [17].
- Complementation Strains: Construct complementation strains by reintroducing the wild-type gene on a plasmid with an inducible promoter (e.g., PBAD) to confirm phenotype specificity [1].
- Mutant Library: Create a panel of single (Δpel, Δpsl), double (ΔpelΔpsl), and hyper-producing (ΔwspF) mutants in the same parental background (e.g., PAO1) for comparative analysis [63] [17].
4.2.2 Phenotypic Characterization of Biofilms
- Static Biofilm Assay (Microtiter Dish): Grow cultures in 96-well plates, stain adhered biomass with crystal violet, and elute for quantitative spectrophotometric measurement (OD570) [17]. This assay is ideal for initial attachment and total biomass quantification.
- Flow-Cell Biofilm & CLSM: Cultivate biofilms under continuous medium flow in flow-cell chambers. Use constitutively GFP-expressing bacteria and specific polysaccharide lectins/antibodies (e.g., Psl-specific lectin) for staining. Image using Confocal Laser Scanning Microscopy (CLSM) and analyze 3D architecture (biovolume, thickness, roughness) with software like COMSTAT [1] [17].
- Quartz Crystal Microbalance with Dissipation (QCM-D): Monitor biofilm formation in real-time on sensor chips. Track changes in resonance frequency (Δf, related to mass) and energy dissipation (ΔD, related to viscoelasticity) to assess the mechanical properties of biofilms formed by different mutants [14].
- Scanning Electron Microscopy (SEM): Fix biofilms, dehydrate using a graded ethanol series, critical-point dry, and sputter-coat with gold/palladium. Image to visualize ultrastructural details of the biofilm matrix and cell morphology [14].
4.2.3 Transcriptomic and Metabolomic Profiling
- RNA Sequencing (RNA-seq): Extract total RNA from planktonic and biofilm cells at mid-exponential phase. Prepare libraries and sequence. Analyze differential gene expression of EPS operons and global regulators in mutant strains compared to wild-type [14].
- Metabolomic Analysis: Quench metabolism, extract intracellular metabolites, and analyze using LC-MS/MS platforms. Identify significant changes in metabolic pathways, particularly in nucleotide sugar precursors (e.g., GDP-mannose for Psl) and central carbon metabolism [14].
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for Investigating P. aeruginosa EPS
| Reagent / Tool | Function / Target | Key Application in Research |
|---|---|---|
| Isogenic EPS Mutant Strains (e.g., PAO1-ΔpslAB, PAO1-ΔpelF) [14] [17] | Specific elimination of a single EPS type. | Fundamental for assigning function and demonstrating compensation between Pel and Psl. |
| Psl-Specific Lectin | Binds to mannose and galactose residues in Psl [1]. | Visualization and quantification of Psl spatial distribution in biofilms via CLSM. |
| Congo Red Dye | Binds to curated amyloid-like fibers and certain polysaccharides like Pel [1]. | Assessment of Pel production through binding in colony morphology assays. |
| Alginate Lyase | Enzymatically degrades alginate polymer [1] [20]. | Testing the functional role of alginate in biofilm integrity and antibiotic susceptibility. |
| Psl Glycoside Hydrolase | Enzymatically degrades Psl polysaccharide [20] [23]. | Disruption of Psl-dependent biofilms; probing Psl as a therapeutic target. |
| Anti-c-di-GMP Antibody | Detects intracellular c-di-GMP levels. | Investigation of the role of this key secondary messenger in regulating the Pel/Psl switch. |
Therapeutic Implications and Future Directions
Understanding EPS compensation is critical for developing effective anti-biofilm strategies. Targeting a single EPS may be ineffective due to redundant functions, suggesting that combination therapies targeting multiple matrix components are necessary.
- Enzyme-Based Therapy: Using glycoside hydrolases that specifically degrade Psl or alginate lyase in combination with antibiotics has been shown to enhance biofilm disruption and bacterial killing [1] [20] [16].
- Phage Therapy: Certain bacteriophages (e.g., PaStL1, PaStL2) have evolved to use Psl as a receptor and produce depolymerases that degrade this polysaccharide. These phages can prevent surface attachment and, in some cases, disrupt mature biofilms, driving bacteria towards a more antibiotic-sensitive, planktonic lifestyle [23].
- Targeting Regulation: Inhibiting enzymes that synthesize c-di-GMP (diguanylate cyclases) could prevent the hyper-aggregative, high-EPS producer phenotypes often selected during chronic infections [17].
The functional redundancy between Pel and Psl represents a powerful bacterial survival strategy. Future therapeutic development must account for this adaptive capacity, moving beyond single-target approaches towards combination strategies that dismantle the entire biofilm matrix and prevent resistance.
The formation of biofilms on industrial and medical surfaces represents a significant challenge across multiple sectors, leading to biofouling, material degradation, and persistent infections. Pseudomonas aeruginosa has emerged as a model organism for biofilm research due to its environmental ubiquity and sophisticated extracellular polymeric substance (EPS) matrix. The structural integrity and protective function of P. aeruginosa biofilms are largely conferred by three exopolysaccharides: alginate, Psl, and Pel [2] [4]. Understanding the distinct and synergistic roles of these biopolymers provides a critical foundation for developing targeted anti-fouling strategies that disrupt biofilm assembly and maintenance at the molecular level. This whitepaper examines the structural and functional characteristics of these EPS components and outlines experimental approaches for exploiting this knowledge in biomaterial surface engineering.
Structural and Functional Characterization of P. aeruginosa Exopolysaccharides
The biofilm matrix of P. aeruginosa incorporates three principal exopolysaccharides with distinct chemical properties and functional roles in biofilm architecture and maintenance. The quantitative and functional distinctions between these polymers are summarized in Table 1.
Table 1: Comparative Analysis of Major P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Charge Properties | Primary Functional Roles | Contribution to Biofilm Integrity |
|---|---|---|---|---|
| Psl | Pentasaccharide repeating units of D-mannose, L-rhamnose, and D-glucose [2] [4] | Neutral [2] | Initial surface attachment [2], microcolony formation [2], structural scaffold in PAO1 strains [3], biofilm elasticity [8] | Dominant regulator of biofilm thickness and elasticity; ΔpslAB mutants show ~66% reduction in biofilm mass [8] |
| Pel | Partially deacetylated α-1,4-linked N-acetylgalactosamine and galactosamine [9] [2] | Cationic [9] [2] | Cell-to-cell interactions [3], structural scaffold in PA14 strains [3], aminoglycoside protection [3], oxidative stress resistance [64] | Redundant enhancement of biofilm biomass; ΔpelF mutants show ~66% reduction in biofilm formation [8] |
| Alginate | Linear copolymer of D-mannuronic acid and L-guluronic acid [4] | Anionic [4] | Biofilm maturation [7], protection against host defenses [7], characteristic of mucoid CF isolates [4] | Negligible impact on initial biofilm architecture in non-mucoid strains; Δalg8 mutants show no significant difference from WT [8] |
The hierarchical importance of these exopolysaccharides varies between strains and environmental conditions. In the widely studied PAO1 strain, Psl serves as the primary structural determinant, with its deletion causing the most severe biofilm defects, while Pel provides complementary functions including enhanced resistance to antimicrobial agents [3] [8]. The biosynthetic pathways for these exopolysaccharides represent promising targets for anti-fouling interventions, as visualized in Figure 1.
Figure 1: Biosynthetic Pathways and Functional Roles of P. aeruginosa Exopolysaccharides. Key enzymes in Psl (PslA, PslB), Pel (PelA, PelF), and alginate (Alg8, AlgD) biosynthesis are shown with their primary functional outputs in biofilm formation and protection.
Experimental Approaches for EPS-Targeted Anti-Fouling Research
Genetic and Molecular Tools for EPS Characterization
Precise genetic manipulation combined with advanced imaging techniques enables systematic dissection of EPS contributions to biofilm formation:
Mutant Strain Construction: Generate isogenic mutants with deletions in key biosynthetic genes (ΔpslAB, ΔpelF, Δalg8) to isolate individual EPS contributions [8]. Employ markerless gene replacement techniques using pEX100T suicide vectors with appropriate antibiotic resistance cassettes (e.g., gentamicin) followed by Flp recombinase-mediated excision from pFLP2 for clean mutant generation [7].
Complementary Expression Systems: For genetic complementation, clone target genes (e.g., pelF) into broad-host-range vectors such as pBBR1-MCS5 under inducible promoters (e.g., lac promoter) and introduce into mutant strains via conjugation or electroporation [7].
High-Resolution Biofilm Imaging: Employ Confocal Laser Scanning Microscopy (CLSM) with polysaccharide-specific lectins or antibodies conjugated to fluorescent dyes (e.g., fluorescein-labeled Psl-specific lectin) to visualize spatial distribution of EPS components [4]. Combine with SEM to examine ultrastructural features of biofilms formed by mutant strains [8].
Quantitative Assessment of Anti-Fouling Strategies
Robust quantification of biofilm formation and material properties is essential for evaluating anti-fouling efficacy:
Crystal Violet Staining Protocol: Grow biofilms in 96-well plates for 24-48 hours, remove planktonic cells by washing, stain with 0.1% crystal violet for 15 minutes, destain with 30% acetic acid, and measure absorbance at 595 nm [8].
Quartz Crystal Microbalance with Dissipation (QCM-D): Monitor biofilm formation in real-time by measuring frequency shift (Δf) and energy dissipation (ΔD) changes on silica or relevant material surfaces. Calculate biofilm mass, thickness, and viscoelastic properties using appropriate modeling software (e.g., QSense Dind v1.0.0) [8].
Single-Channel Conductivity Measurements: For pore-forming EPS export proteins (e.g., PelB), incorporate purified protein into lipid bilayers and measure ionic current to assess channel gating dynamics and the effect of inhibitory compounds [9].
The quantitative outcomes of these approaches for different EPS mutants are summarized in Table 2.
Table 2: Quantitative Biofilm Parameters for P. aeruginosa EPS Mutants
| Strain | Biofilm Mass (Relative to WT) | Biofilm Thickness | Elastic Modulus | Key Experimental Findings |
|---|---|---|---|---|
| PAO1-WT | 100% | ~12.5 μm [8] | Baseline value | Forms structured biofilms with characteristic mushroom-like microcolonies [7] |
| PAO1-ΔpslAB | ~33% [8] | Significantly decreased [8] | ~4x increase [8] | Severe attachment defect; essential for biofilm maturity and architecture [3] [8] |
| PAO1-ΔpelF | ~33% [8] | Reduced [8] | No significant change [8] | Increased susceptibility to aminoglycosides and oxidative stressors [64] [3] |
| PAO1-Δalg8 | No significant difference [8] | Similar to WT [8] | No significant change [8] | Develops biofilms with decreased viable cells; enhanced eDNA structures [7] |
Targeted Anti-Fouling Strategies and Research Toolkit
Enzymatic and Molecular Interventions
EPS-degrading enzymes represent promising anti-biofilm agents with high specificity:
Glycoside Hydrolases: Psl glycoside hydrolase specifically degrades the Psl polysaccharide backbone, disrupting biofilm integrity and increasing antibiotic penetration [20]. Application of Pslgh in combination with standard antibiotics significantly reduces bacterial load on surfaces and in 3D lung cell aggregates [20].
Alginate Lyase: Breaks the glycosidic linkages in alginate, particularly effective against mucoid strains. When combined with antibiotics, demonstrates synergistic effects in biofilm eradication [2] [20].
Dispersin B: Hydrolyzes PNAG (poly-N-acetylglucosamine) in other species, though not naturally produced by P. aeruginosa, provides a conceptual framework for EPS-degrading enzyme applications [65].
The workflow for developing and testing enzymatic anti-fouling strategies is illustrated in Figure 2.
Figure 2: Workflow for Developing Enzymatic Anti-Fouling Strategies. The process begins with EPS target identification, proceeds through enzyme selection and assay development, and culminates in combination testing with conventional antimicrobials or surface modifications.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Essential Research Reagents for EPS-Targeted Anti-Fouling Research
| Reagent/Material | Specific Examples | Research Application | Functional Role |
|---|---|---|---|
| Genetic Tools | pEX100T suicide vector, pBBR1-MCS5 expression vector, pFLP2 recombinase plasmid [7] | Mutant construction and genetic complementation | Targeted gene deletion and controlled gene expression for functional studies |
| Detection Reagents | Fluorescein-conjugated Psl-specific lectin, Congo red, Crystal violet [3] [8] | EPS visualization and quantification | Specific staining and quantification of exopolysaccharides in biofilms |
| Enzymes | Psl glycoside hydrolase, Alginate lyase [20] | EPS degradation studies | Targeted disruption of specific exopolysaccharides to compromise biofilm integrity |
| Analytical Systems | Quartz Crystal Microbalance with Dissipation (QCM-D) sensors, Confocal Laser Scanning Microscope [8] | Real-time biofilm monitoring | Label-free, real-time assessment of biofilm formation and material properties |
| Model Surfaces | Silica chips, Polycarbonate membranes, 3D lung epithelial cell aggregates [20] [8] | Anti-fouling surface testing | Standardized substrates for evaluating biofilm formation and anti-fouling efficacy |
The targeted disruption of exopolysaccharide systems in P. aeruginosa represents a promising approach for next-generation anti-fouling surface technologies. The distinct yet complementary roles of Psl, Pel, and alginate in biofilm formation and maintenance enable strategic interventions that address specific fouling challenges across industrial and medical applications. Future research directions should focus on the development of specific Pel-degrading enzymes, which currently represent a significant gap in the anti-fouling toolkit, and the engineering of surface coatings that incorporate synergistic combinations of EPS-targeting agents. As structural insights into EPS export machinery deepen [9] and multi-omics approaches clarify regulatory networks [8], increasingly sophisticated anti-fouling strategies will emerge to address the persistent challenge of biofilm-mediated fouling and resistance.
Evidence and Efficacy: Validating the Unique and Synergistic Contributions of Each EPS
The biofilm matrix formed by Pseudomonas aeruginosa constitutes a critical virulence determinant, providing structural integrity and protection in diverse environments, including the cystic fibrosis (CF) lung. This extracellular matrix is a complex mixture of polymers, with exopolysaccharides serving as a primary scaffold. Alginate, Pel, and Psl are the three major exopolysaccharides implicated in biofilm development, yet their functional preeminence varies significantly across different P. aeruginosa strains [17] [16]. Understanding which polysaccharide serves as the dominant structural scaffold is not merely academic; it has profound implications for designing targeted anti-biofilm therapies. This review synthesizes evidence demonstrating that the structural superiority of Pel versus Psl is not absolute but is fundamentally strain-dependent, with functional redundancy providing an adaptive safeguard for biofilm integrity [17] [45] [3].
Functional Roles of Pel and Psl in the Biofilm Matrix
While both Pel and Psl contribute to biofilm formation, they exhibit distinct functional specializations and chemical properties that influence their efficacy as structural scaffolds.
Psl is a neutral, branched pentasaccharide composed of repeating D-mannose, D-glucose, and L-rhamnose subunits [17] [16]. It plays a critical role in the initial stages of biofilm development, serving as a primary adhesin for surface attachment [17] [3]. As the biofilm matures, Psl provides structural integrity by cross-linking with the adhesin CdrA and extracellular DNA (eDNA) [16]. It also functions as a scaffold for the development of the mature biofilm structure and is crucial for cell-cell interactions and surface adhesion [17].
Pel is a cationic polymer composed of partially de-N-acetylated α-1,4-N-acetylgalactosamine, consisting predominantly of galactosamine and N-acetylgalactosamine repeats [16] [66]. Its positive charge enables it to cross-link with negatively charged eDNA, facilitating cell-to-cell interactions and enhancing the structural stability of the biofilm community [16] [3]. Pel is particularly important for pellicle formation at the air-liquid interface and contributes significantly to the later stages of biofilm maturation [66].
Table 1: Comparative Functional Profiles of Pel and Psl Polysaccharides
| Feature | Pel Polysaccharide | Psl Polysaccharide |
|---|---|---|
| Chemical Nature | Cationic polymer [16] | Neutral, branched pentasaccharide [17] [16] |
| Primary Role | Cell-cell interactions, biofilm maturation [17] [3] | Initial surface attachment, structural scaffold [17] [16] |
| Charge Characteristics | Positively charged [16] | Neutral [16] |
| Key Interactions | Cross-links with negatively charged eDNA [16] | Cross-links with CdrA adhesin and eDNA [16] |
| Antibiotic Protection | Protection against aminoglycosides [3] | Limited penetrance barrier function [16] |
Strain-Dependent Variation in Polysaccharide Dependency
The structural reliance on either Pel or Psl is not uniform across the P. aeruginosa species but demonstrates marked strain-to-strain variability. This was systematically investigated through mutational analysis of various clinical and environmental isolates, which revealed distinct functional classes based on Pel and Psl dependency profiles [17].
The paradigm for this strain-specificity is illustrated by the two most commonly studied laboratory strains:
- PAO1: Utilizes Psl as the primary structural polysaccharide. Psl is critical for both initial attachment and mature biofilm structure in this strain. A psl mutant in PAO1 is severely deficient in attachment and arrested at the monolayer stage in flow-cell biofilms [17] [3].
- PA14: Relies predominantly on Pel for biofilm integrity. PA14 carries a natural deletion in part of the psl operon and consequently depends on Pel for maintaining cell-to-cell interactions and mature biofilm architecture. A pel mutant in PAO1 has no observable phenotype, whereas in PA14, it results in a severe biofilm deficiency [17] [3].
This strain-specific dependency extends beyond laboratory strains. Analysis of clinical and environmental isolates revealed that while a psl mutation consistently impaired initial surface attachment across all strains tested, the contribution of Pel and Psl to mature biofilm structure was highly variable [17]. This led to the proposal of four distinct classes of P. aeruginosa strains based on their Pel and Psl functional and expression profiles, underscoring the genetic diversity influencing matrix composition [17].
Quantitative Analysis of Mutant Phenotypes
The structural roles of Pel and Psl have been quantified using various biofilm assays, revealing clear strain-specific dependencies. The data below summarize findings from microtiter dish assays and flow-cell reactor experiments that measure biofilm biomass and structure in wild-type and mutant strains [17] [3].
Table 2: Quantitative Biofilm Phenotypes of P. aeruginosa Strains and Mutants
| Strain | Genotype | Surface Attachment | Mature Biofilm Biomass | Biofilm Architecture |
|---|---|---|---|---|
| PA14 | Wild-type | Normal [3] | High [17] [3] | Normal, complex structures [17] |
| PA14 | pel mutant | Normal [3] | Substantially reduced [17] [3] | Arrested at monolayer [17] |
| PA14 | psl mutant | Normal (operon deleted) [17] | High [17] | Normal, complex structures [17] |
| PAO1 | Wild-type | Normal [3] | High [17] [3] | Normal, complex structures [17] |
| PAO1 | pel mutant | Normal [3] | Normal [17] [3] | Normal, complex structures [17] |
| PAO1 | psl mutant | Severely deficient [17] [3] | Impaired [17] [3] | Arrested at monolayer [17] |
| PAO1 | pel psl double mutant | Severely deficient [17] | Impaired [17] | Arrested at monolayer [17] |
Regulatory Control and Functional Redundancy
The expression of Pel and Psl is governed by a complex regulatory network that ensures appropriate production under varying environmental conditions.
Key Regulatory Pathways
A central regulator is FleQ, which acts as a repressor for both the pel and psl operons. This repression is relieved when FleQ binds the secondary messenger cyclic-di-GMP (c-di-GMP) [17]. Elevated intracellular levels of c-di-GMP are a hallmark of the rugose small colony variant (RSCV) phenotype, which is associated with hyper-adherence and hyper-aggregation due to increased production of Pel and Psl [17]. Furthermore, c-di-GMP can allosterically activate Pel synthesis through direct binding to PelD [17]. Other important regulators include:
- RpoS: Acts as a positive transcriptional regulator of psl [17].
- Quorum Sensing: Positively influences both pel and psl expression [17].
- Gac-Rsm Pathway: The RNA-binding protein RsmA inhibits psl translation, providing post-transcriptional control [17].
This intricate regulation allows P. aeruginosa to fine-tune polysaccharide production in response to environmental cues.
Functional Redundancy and Evolvability
The capacity to produce multiple exopolysaccharides provides functional redundancy, which serves as an evolutionary safeguard for biofilm formation. This was demonstrated through a compelling evolution experiment with PAO1, a strain that primarily utilizes Psl. When a PAO1 psl mutant, which initially produced poor biofilms, was subjected to extended cultivation, it acquired compensatory mutations that upregulated Pel expression [17] [45]. This adaptation allowed the population to regain the capacity to form robust biofilms, highlighting the utility of possessing a redundant scaffold exopolysaccharide. This redundancy ensures that even if one polysaccharide system is compromised, the other can be co-opted to preserve the biofilm lifestyle, a key survival advantage [17] [45].
Diagram: Regulatory interplay and functional redundancy in Pel and Psl production. Elevated c-di-GMP levels, triggered by environmental cues, activate FleQ, which in turn promotes the expression of both the pel and psl operons. The resulting production of Pel and Psl polysaccharides facilitates mature biofilm formation. The dashed green line highlights the critical phenomenon of functional redundancy, wherein one polysaccharide can compensate for the loss of the other.
Experimental Approaches for Polysaccharide Analysis
Key Methodologies
Research elucidating the roles of Pel and Psl relies on a suite of established biochemical and microbiological techniques.
Static Microtiter Dish Assay: This method quantifies biofilm biomass that forms at the air-liquid interface after static incubation. Bacterial strains are inoculated into the wells of a microtiter plate and incubated without agitation. Non-adherent cells are removed by washing, and the adherent biofilm biomass is quantified using stains like crystal violet or through optical density measurements [17] [3]. This assay is particularly useful for high-throughput screening of attachment and biofilm formation capabilities.
Flow-Cell Reactor with Confocal Microscopy: This system allows for the visual monitoring of live biofilms growing under continuous flow of dilute medium. It provides a dynamic environment that more closely mimics natural conditions. Biofilms grown in flow cells are typically analyzed using Confocal Laser Scanning Microscopy (CLSM), which generates high-resolution, three-dimensional images of the biofilm architecture without disrupting the native structure [17]. This technique is essential for visualizing developmental stages, from initial attachment to mature microcolonies.
Genetic Mutagenesis and Complementation: A common strategy involves generating in-frame deletion mutants of key biosynthetic genes, such as pelF for Pel and pslD for Psl, to abolish the production of the respective polysaccharide [17]. The phenotypes of these mutants are then compared to the wild-type strain in the assays described above. Complementing the mutation by reintroducing the functional gene on a plasmid is crucial to confirm that the observed phenotype is directly linked to the loss of the polysaccharide and not to secondary mutations.
Essential Research Reagents
The following table catalogues critical reagents and their applications for studying P. aeruginosa biofilm polysaccharides.
Table 3: Research Reagent Solutions for Biofilm Polysaccharide Studies
| Reagent / Material | Function / Application | Experimental Context |
|---|---|---|
| Congo Red Dye | Binds to polysaccharides; used to assess Pel production via colony staining and liquid binding assays [3]. | Phenotypic screening of polysaccharide production [3]. |
| Arabinose-Inducible System (P_BAD) | Controls conditional gene expression; used to create Pel overexpression strains for functional analysis [3]. | Genetic complementation and overexpression studies [3]. |
| Crystal Violet Stain | Stains adhered cells and matrix components; quantifies total biofilm biomass in microtiter plate assays [66]. | High-throughput quantification of biofilm formation [66]. |
| Fluorescent Protein Tags (e.g., GFP, mCherry) | Labels specific bacterial strains for visualization; allows distinction between different populations in co-culture or recolonization experiments [6]. | Confocal microscopy and spatial analysis in flow cells [6]. |
| N-Acetylcysteine (NAC) | Matrix-penetrating antimicrobial that kills biofilm cells while leaving the EPS matrix intact for mechanical studies [6]. | Studying the physical properties of the residual matrix [6]. |
The determination of Pel versus Psl as the primary structural scaffold in P. aeruginosa is a definitive example of strain-specific adaptation. Neither polysaccharide is universally superior; rather, their structural preeminence depends on the genetic background of the strain, with PAO1 and PA14 serving as canonical examples of Psl- and Pel-dependency, respectively. This variability is managed by a sophisticated regulatory network, with c-di-GMP serving as a key integrator of environmental signals to direct polysaccharide production. Critically, the functional redundancy between Pel and Psl provides a robust evolutionary strategy for preserving biofilm integrity, as evidenced by the compensatory upregulation of Pel in Psl-deficient mutants. Future therapeutic strategies aimed at disrupting the biofilm matrix must account for this strain-level variation and functional redundancy, potentially requiring multi-target approaches or diagnostics that profile a strain's exopolysaccharide expression signature prior to treatment.
Pseudomonas aeruginosa is a formidable opportunistic pathogen whose resilience in both environmental and clinical settings is largely dictated by its biofilm mode of growth. The extracellular matrix of these biofilms, a complex scaffold of polymers, provides critical protection against antimicrobial agents and host defenses. While alginate has been extensively studied in chronic infections, the exopolysaccharides Pel and Psl are increasingly recognized for their pivotal roles in the biofilm architecture of non-mucoid strains. This whitepaper synthesizes current research to validate the specific functions of Pel and Psl as integral components of a protective shield, with a focused analysis of their direct contributions to oxidative stress resistance and antibiotic tolerance. Framed within the broader context of P. aeruginosa biofilm matrix research, we present a detailed examination of their biosynthetic pathways, protective mechanisms, and the experimental methodologies that underpin these findings, providing a technical resource for researchers and drug development professionals.
The biofilm matrix of Pseudomonas aeruginosa is a key determinant of its success as a versatile and persistent bacterium. This matrix is a composite of extracellular polymeric substances (EPS), including nucleic acids, proteins, lipids, and polysaccharides. Among these, the exopolysaccharides alginate, Pel, and Psl are critical for biofilm structure and function [4]. Alginate production is often associated with mucoid variants isolated from the chronic infections that afflict individuals with cystic fibrosis (CF). In contrast, most environmental and clinical isolates are non-mucoid and rely primarily on Pel and Psl for biofilm development [17] [4]. These two polysaccharides are not merely structural; they are active mediators of the community's ability to withstand environmental insults. This technical guide delves into the specific roles of Pel and Psl, moving beyond their structural contributions to highlight their validated functions as a protective shield against two major classes of threats: reactive oxygen species (from both host immune responses and environmental sources) and a broad spectrum of antibiotics. Understanding these mechanisms is paramount for developing novel strategies to disrupt biofilm-associated resistance.
Pel and Psl are chemically distinct exopolysaccharides synthesized by dedicated machinery, which dictates their unique physical properties and functional roles in the biofilm matrix.
Pel: A Cationic Structural Scaffold
- Chemical Structure: Pel is a linear polymer of α-1,4-linked N-acetylgalactosamine (GalNAc) that is partially de-N-acetylated during its biosynthesis [5]. This de-N-acetylation introduces positively charged galactosamine (GalN) residues, rendering the overall polymer cationic at physiological pH.
- Biosynthetic Machinery: The
pelABCDEFGoperon encodes the proteins responsible for Pel synthesis, modification, and export. The proposed biosynthetic model shares features with the alginate pathway. The proteins form a complex that spans the cellular envelope: PelF is a cytoplasmic glycosyltransferase, PelD and PelE form an inner membrane complex, PelA is a periplasmic deacetylase, and PelB and PelC form the outer membrane export channel [9] [4]. - Primary Functions: Pel is crucial for maintaining cell-to-cell interactions and providing structural integrity to mature biofilms, particularly in pellicles (biofilms at the air-liquid interface) [3]. Its cationic nature facilitates electrostatic interactions with anionic components of the matrix, such as extracellular DNA, thereby cross-linking and stabilizing the biofilm structure [5].
Psl: A Neutral Adhesive Framework
- Chemical Structure: Psl is a neutral polysaccharide composed of a repeating pentasaccharide unit containing D-mannose, L-rhamnose, and D-glucose [4]. Its biosynthesis resembles the Wzy-dependent pathway used for capsular polysaccharide synthesis in E. coli, involving the assembly of subunits on an isoprenoid lipid carrier [4].
- Biosynthetic Machinery: The
psloperon (e.g.,pslAthroughpslOin PAO1) encodes the enzymes for monosaccharide synthesis, assembly, and export. PslB, for instance, is a bifunctional enzyme with phosphomannose isomerase (PMI) and GDP-mannose pyrophosphorylase (GMP) activities, crucial for synthesizing sugar-nucleotide precursors [67]. - Primary Functions: Psl plays a dominant role in the initial attachment of cells to both biotic and abiotic surfaces [17] [18]. It forms a fiber-like matrix that surrounds cells, facilitating cell-surface and cell-cell interactions and is essential for the structural stability of biofilms in many strains [4].
Table 1: Core Characteristics of Pel and Psl Polysaccharides
| Feature | Pel Polysaccharide | Psl Polysaccharide |
|---|---|---|
| Chemical Nature | Cationic, linear polymer | Neutral, branched polymer |
| Monosaccharide Composition | α-1,4-linked GalNAc/GalN [5] | D-mannose, D-glucose, L-rhamnose [4] |
| Primary Biosynthetic Locus | pelABCDEFG operon [3] |
psl operon (e.g., pslA-O) [67] |
| Key Structural Role | Cell-cell interactions, pellicle formation, structural scaffold [3] | Initial surface attachment, biofilm structural stability [18] |
| Charge at Physiological pH | Positive [5] | Neutral |
Protective Role Against Oxidative Stress
Exposure to reactive oxygen species (ROS) is a common stressor for P. aeruginosa, originating from environmental sources like solar UVA radiation or from the host immune response during infection. Recent research demonstrates that Pel and Psl are critical for adaptive responses and survival under such oxidative duress.
Response to UVA Radiation
Sublethal UVA radiation induces oxidative damage and triggers adaptive responses in P. aeruginosa. A 2023 study demonstrated that this exposure upregulates the expression of pelA and pslA genes, leading to increased polysaccharide production [66]. This response is functional, as the study further showed that the presence of Pel and Psl was essential for the UVA-induced increase in:
- Cell hydrophobicity
- Cell attachment capability
- Cross-protection against subsequent lethal doses of UVA, hydrogen peroxide (H₂O₂), and sodium hypochlorite [66]
Furthermore, Pel and Psl were critical for the direct tolerance of planktonic cells and biofilms to lethal doses of these oxidizing agents. Mutants unable to produce both Pel and Psl (∆pel psl) showed the most severe survival defects, underscoring a potentially synergistic role in oxidative stress defense [66].
Defense against Neutrophil-Generated Oxidants
Within a host, neutrophils attack bacteria with a cocktail of ROS, including H₂O₂ and hypochlorous acid (the active component of sodium hypochlorite). The capacity of Pel and Psl to provide tolerance to H₂O₂ and sodium hypochlorite, as validated in the UVA study, directly translates to enhanced survival against this arm of the innate immune response [66]. Murakami et al. (cited in [66]) previously reported that stationary-phase cultures of a psl mutant have lower tolerance to H₂O₂, reinforcing the role of Psl in antioxidative defense.
Role in Antibiotic Resistance
Biofilms are notorious for their inherent tolerance to antibiotics, and Pel and Psl are significant contributors to this phenotype through both physical and physiological mechanisms.
Pel and Aminoglycoside Tolerance
Research by Colvin et al. established that Pel plays a specific and direct role in enhancing tolerance to aminoglycoside antibiotics in biofilm populations. This protection was not observed in planktonic cells, indicating a biofilm-specific mechanism. The cationic nature of Pel, resulting from its de-N-acetylated residues, is hypothesized to function as a cationic scavenger, binding and sequestering the positively charged aminoglycoside molecules (e.g., tobramycin, gentamicin), thereby reducing their effective concentration at the cell surface [3] [5].
Psl as a Broad-Spectrum Barrier
Psl contributes to antibiotic resistance through multiple mechanisms. It acts as a physical diffusion barrier, hindering the penetration of antimicrobial agents through the biofilm matrix [68]. A clinical study of 100 isolates found a significant statistical relationship (p < 0.05) between the presence of the pslB gene and multidrug resistance (MDR) in high-biofilm-forming strains [67]. Furthermore, Psl has been shown to interact with other bacterial species in polymicrobial biofilms, such as Staphylococcus aureus, increasing P. aeruginosa's resistance to antibiotics [66] [68].
Table 2: Documented Roles of Pel and Psl in Stress Resistance
| Stress Type | Polysaccharide | Protective Mechanism & Key Evidence |
|---|---|---|
| UVA Radiation | Pel and Psl | Sublethal UVA induces pelA/pslA expression; essential for cross-protection and direct tolerance to lethal UVA, H₂O₂, and NaOCl [66]. |
| H₂O₂ (Oxidative Stress) | Psl | Stationary-phase psl mutants show lower H₂O₂ tolerance [66]. |
| Pel and Psl | Simultaneous presence is key for tolerance in planktonic and biofilm states [66]. | |
| Aminoglycosides | Pel | Provides biofilm-specific tolerance; cationic polymer may sequester cationic antibiotics [3]. |
| Multidrug Resistance | Psl | Significant association (p < 0.05) between pslB gene presence and MDR in high-biofilm-forming clinical isolates [67]. |
Essential Experimental Protocols and Methodologies
Validating the roles of Pel and Psl requires a toolkit of well-established molecular microbiology and biochemical assays. Below are detailed protocols for key experiments cited in this field.
Genetic Manipulation: Generating Polysaccharide-Deficient Mutants
The foundational step for functional studies is the creation of isogenic mutants with targeted deletions in Pel or Psl biosynthesis genes.
- Objective: To construct in-frame, markerless deletion mutants (e.g., ∆pelA, ∆pslD) in a specific P. aeruginosa background (e.g., PAO1).
- Procedure:
- Fragment Amplification: Using polymerase chain reaction (PCR), amplify ~500-1000 bp DNA fragments upstream and downstream of the target gene (e.g.,
pelForpslD). - Vector Construction: Ligate these fragments into a suicide vector (e.g., pEX18Gm) that cannot replicate in P. aeruginosa and contains a counterselectable marker (e.g., sacB).
- Conjugation: Introduce the constructed plasmid into P. aeruginosa via conjugation with an E. coli donor strain.
- Selection and Counter-Selection: Select for single-crossover integrants using antibiotic resistance. Subsequently, plate integrants on media containing sucrose to select for a second crossover event, which excises the plasmid and may result in the desired gene deletion.
- Verification: Confirm the deletion by colony PCR and sequencing of the genomic locus [17].
- Fragment Amplification: Using polymerase chain reaction (PCR), amplify ~500-1000 bp DNA fragments upstream and downstream of the target gene (e.g.,
Biofilm Phenotyping: Static Microtiter Dish Assay
This high-throughput method quantifies biofilm formation capacity.
- Objective: To assess and quantify the biofilm-forming ability of wild-type and mutant strains.
- Procedure:
- Inoculation: Grow bacterial cultures to mid-exponential phase, dilute to a standardized optical density (e.g., OD₆₅₀ ~0.1), and aliquot 100-200 µL into the wells of a sterile 96-well polystyrene microtiter plate.
- Incubation: Incubate the plate statically for a defined period (e.g., 16-48 hours) at the desired temperature (e.g., 37°C).
- Washing: Carefully aspirate the planktonic cells and rinse the wells with a saline solution (e.g., phosphate-buffered saline) to remove non-adherent bacteria.
- Staining and Elution: Fix the adhered biofilm with methanol and stain with a 1% crystal violet solution for 15-20 minutes. Wash away excess stain, then solubilize the bound crystal violet with a 33% glacial acetic acid solution.
- Quantification: Measure the optical density of the eluted dye at 570-600 nm using a microplate reader. Higher OD values correlate with greater biofilm biomass [67] [3].
Assessing Antibiotic and Oxidant Tolerance
- Objective: To determine the minimum biofilm eradication concentration (MBEC) or survival of biofilms after antimicrobial challenge.
- Procedure:
- Biofilm Growth: Grow biofilms in a standardized system (e.g., Calgary Biofilm Device, flow cells, or 96-well plates).
- Challenge: Expose mature biofilms to a range of concentrations of the target antibiotic (e.g., tobramycin) or oxidizing agent (e.g., H₂O₂, NaOCl) for a specified time.
- Viability Assessment: For oxidant stress, cells may be harvested immediately after exposure. For antibiotic treatment, biofilms are often disrupted, and the bacterial suspension is serially diluted and spot-plated on non-selective agar to enumerate surviving colony-forming units (CFUs) after a period of recovery [66] [3].
Visualization of Biosynthesis and Experimental Logic
The following diagrams, generated using DOT language, illustrate the core biosynthetic pathway of Pel and the experimental logic used to dissect its function.
Pel Biosynthesis and Export Pathway
Experimental Workflow for Functional Validation
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents and Strains for Investigating Pel/Psl Function
| Reagent / Material | Function / Application | Key Examples / Notes |
|---|---|---|
| Defined Bacterial Strains | Provides isogenic background for functional comparisons. | PAO1 (relies on Psl), PA14 (relies on Pel), PAO1∆pel, PAO1∆psl, PAO1∆pel psl [66] [17]. |
| Suicide Vectors | Enables construction of markerless, in-frame gene deletions. | pEX18Gm (or similar), contains sacB for counter-selection [17]. |
| Microtiter Plates | High-throughput quantification of biofilm formation. | 96-well polystyrene plates for crystal violet assay [67]. |
| Flow Cell Reactors | Study biofilm development under shear stress with microscopy. | Allows for real-time, non-destructive imaging of biofilm architecture [17] [3]. |
| Specific Lectins | Visualize and localize polysaccharides in biofilms. | Wisteria floribunda lectin (WFL) binds Pel [5]. |
| Glycoside Hydrolases | Selectively degrade polysaccharides for structural/functional studies. | PelAh (hydrolase domain of PelA) cleaves Pel polymer [5]. |
| c-di-GMP Modulators | Manipulate intracellular c-di-GMP levels to induce polysaccharide production. | High c-di-GMP levels (e.g., via ∆wspF mutation) upregulate Pel/Psl [5]. |
The body of evidence unequivocally validates Pel and Psl as critical components of P. aeruginosa's "protective shield." Beyond their established roles in biofilm architecture, these exopolysaccharides are active mediators of resistance against oxidative stress and a diverse array of antibiotics. The cationic property of Pel facilitates specific interactions that sequester aminoglycosides, while Psl provides a robust physical barrier and is clinically correlated with multidrug resistance. Their induction in response to sublethal UVA radiation further highlights their function as dynamic, adaptive response elements.
Future research should focus on exploiting these vulnerabilities. Strategies that inhibit Pel and Psl biosynthesis, disrupt their export, or enzymatically degrade these polymers (e.g., using PelAh) represent promising therapeutic avenues. Combining such matrix-disrupting agents with conventional antibiotics could potentially break the biofilm barrier, resensitizing persistent infections to treatment. As part of the broader alginate-Pel-Psl matrix system, understanding the regulatory interplay and functional redundancy between these polysaccharides will be crucial for developing effective anti-biofilm strategies to combat resilient P. aeruginosa infections.
The extracellular matrix of Pseudomonas aeruginosa biofilms represents a critical determinant of the pathogen's resilience in chronic infections, such as those occurring in cystic fibrosis lungs and chronic wounds [2] [69]. This matrix functions not as a mere physical barrier but as a dynamically interactive system wherein the major components—alginate, Psl, Pel, and extracellular DNA (eDNA)—engage in specific molecular interactions that collectively enhance structural integrity and protective function [69]. Understanding the precise nature of these synergistic interactions is paramount for developing effective therapeutic strategies against biofilm-mediated infections [2]. This technical review examines the cross-linking mechanisms that fortify the P. aeruginosa biofilm matrix, integrating current structural insights with experimental approaches for investigating matrix properties.
Molecular Profiles of Matrix Components
The P. aeruginosa biofilm matrix consists of a complex network of biopolymers with diverse chemical properties that enable specific inter-molecular interactions. The table below summarizes the key characteristics and primary functions of each major matrix component.
Table 1: Molecular characteristics and functions of major matrix components in P. aeruginosa biofilms
| Component | Chemical Nature | Monomeric Composition | Primary Functions in Matrix |
|---|---|---|---|
| Alginate | Anionic, acetylated linear polymer [69] | β-D-mannuronic acid and α-L-guluronic acid [8] | Matrix maturation, viscoelasticity, antibiotic diffusion limitation, immune evasion [69] |
| Psl | Neutral branched pentasaccharide [2] [69] | D-mannose, D-glucose, and L-rhamnose [2] [8] | Surface attachment, cell-cell interactions, structural stability, signaling via c-di-GMP [69] |
| Pel | Cationic, partially deacetylated polymer [2] [63] | N-acetylgalactosamine and N-acetylglucosamine [2] [8] | Pellicle formation, cell-cell adherence, aminoglycoside tolerance via charge-based interactions [63] [69] |
| eDNA | Poly-anionic double-stranded polymer [69] | Deoxyribonucleotides | Cation chelation, cell organization, acidification of biofilm environment, neutrophil activation [69] |
Cross-Linking Mechanisms in the Matrix Network
Charge-Based Interactions Between Pel and eDNA
The cationic nature of Pel polysaccharide enables direct electrostatic interaction with the negatively charged phosphate backbone of eDNA, forming a fundamental cross-linking mechanism within the biofilm matrix [2]. This ionic bonding creates a stable network that contributes significantly to structural integrity and promotes bacterial aggregation [2] [63]. The functional consequence of this interaction is enhanced mechanical stability and protection from aminoglycoside antibiotics, which are positively charged and may be sequestered within the Pel-eDNA network [2] [69]. Furthermore, this association protects eDNA from degradation by DNase I, potentially rendering nuclease-based therapies less effective [2].
Cationic Bridging Mediated by Divalent Cations
Divalent cations, particularly calcium (Ca²⁺), serve as crucial intermediaries in matrix cross-linking through cationic bridging [60]. In this mechanism, Ca²⁺ ions form simultaneous coordinate bonds with multiple anionic components, creating extensive inter-polymer networks [60]. This bridging occurs between guluronic acid residues in alginate chains (forming the characteristic "egg-box" structures), between alginate and eDNA, and between different eDNA molecules [6] [60]. The resulting network significantly increases matrix density and reduces permeability to antimicrobial agents [60].
Alginate-Driven Donnan Effect and Matrix Swelling
The high density of negatively charged carboxyl groups on alginate polymers creates a polyelectrolyte environment that exerts substantial influence on matrix physical properties through the Donnan effect [6]. This effect generates an osmotic pressure gradient that drives water influx into the matrix, resulting in pronounced swelling behavior [6]. The swollen alginate-rich matrix exhibits enhanced elastic modulus and mechanical stability while maintaining hydrogel permeability for nutrient diffusion [6]. This swelling phenomenon is particularly characteristic of mucoid P. aeruginosa strains that overproduce alginate [6].
Structural Redundancy and Complementary Functions
The matrix network exhibits functional redundancy between Psl and Pel polysaccharides, which can compensate for each other structurally despite their differing chemical properties [63] [8]. While Psl dominates in maintaining biofilm elasticity and thickness in strains like PAO1, Pel can enhance biofilm biomass and assume structural roles when Psl is absent [8]. This redundancy provides resilience to the matrix against enzymatic targeting and ensures structural stability across varying environmental conditions [63].
Figure 1: Cross-linking mechanisms between matrix components and their functional consequences in P. aeruginosa biofilms
Experimental Approaches for Investigating Matrix Interactions
Methodologies for Structural Analysis
Advanced structural biology techniques have been instrumental in elucidating the molecular architecture of exopolysaccharide export and assembly mechanisms.
Cryo-Electron Microscopy of Pel Export Complex: The PelBC export complex responsible for Pel exopolysaccharide translocation across the outer membrane has been characterized using single-particle cryo-EM [9]. The experimental protocol involves:
- Heterologous expression: PelB (residues 762-1193) and PelC with modified signal sequences are expressed in E. coli C41(DE3) ΔompF ΔacrAB strain [9].
- Membrane extraction and purification: Proteins are extracted with n-dodecyl-β-D-maltoside (DDM) detergent, followed by immobilized metal affinity chromatography (IMAC) exploiting PelB's histidine tag [9].
- Nanodisc reconstitution: The complex is stabilized in POPC:POPG (70:30) lipid nanodiscs using MSP1D1 scaffold protein [9].
- Cryo-EM imaging: Data collection from ~30,000 movies yielding 4.8 million particles followed by 3D reconstruction at 2.5 Å resolution [9].
This approach revealed the 1:12 stoichiometry of the PelB:PelC complex and identified critical protein-lipid interactions stabilizing the assembly [9].
Molecular Dynamics Simulations of Gating Mechanisms: Computational approaches complement structural data by providing dynamic insights:
- All-atom molecular dynamics simulations have been employed to study the gating mechanism of the PelB β-barrel, revealing the flexibility of the extracellular Plug-S loop that facilitates polysaccharide transport [9].
- Simulations of alginate oligosaccharide interactions with Ca²⁺ and DNA have demonstrated the disruption of cationic bridging within the matrix network [60].
Methodologies for Functional Characterization
Functional assays quantify the mechanical and protective properties arising from matrix component interactions.
Particle-Tracking Microrheology (PTM): This technique measures localized viscoelastic properties within intact biofilms [6]:
- Sample preparation: Fluorescent microparticles (0.5-1.0 μm) are incorporated during biofilm formation or gently infused into pre-formed biofilms [6].
- Image acquisition: Time-lapse imaging tracks Brownian motion of particles within biofilm microcolonies using confocal laser scanning microscopy [6].
- Data analysis: Mean square displacement (MSD) is calculated from particle trajectories and converted to viscoelastic moduli (G' for elastic storage modulus and G" for viscous loss modulus) [6].
PTM has demonstrated that alginate-overproducing biofilms exhibit significantly higher elastic modulus and swelling capacity compared to Psl-dominated matrices [6].
COMSTAT Analysis of Biofilm Architecture: Quantitative image analysis software enables precise measurement of biofilm structural parameters:
- Biofilm staining: Matrix components are specifically labeled - alginate with Texas Red-labeled oligomers, eDNA with TOTO-1 or SYTO dyes, proteins with FITC conjugation [60].
- Confocal imaging: z-stack images are collected at multiple random positions within each sample [60].
- Image analysis: COMSTAT software quantifies biovolume (μm³/μm²), average thickness (μm), surface area coverage, and roughness coefficient [60].
This approach has revealed that OligoG treatment (≥2%) significantly reduces EPS polysaccharides and eDNA content in established biofilms [60].
Table 2: Experimental techniques for investigating matrix interactions and properties
| Technique | Measured Parameters | Key Applications in Matrix Research |
|---|---|---|
| Cryo-EM | Molecular structure, protein-lipid interactions, complex stoichiometry | Visualization of Pel export machinery [9] |
| Particle-Tracking Microrheology | Local viscoelastic moduli (G', G"), mesh size, permeability | Spatial mapping of mechanical heterogeneity in alginate-rich vs Psl-rich biofilms [6] |
| COMSTAT Analysis | Biofilm biovolume, thickness, roughness, surface coverage | Quantifying structural disruption after matrix-targeting treatments [60] |
| Quartz Crystal Microbalance with Dissipation (QCM-D) | Mass adsorption, thickness, viscoelastic properties in real-time | Monitoring biofilm development kinetics and response to anti-biofilm agents [8] |
| Molecular Dynamics Simulations | Molecular interactions, binding energies, conformational dynamics | Predicting disruption of Ca²⁺ bridging by alginate oligosaccharides [60] |
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential research reagents and materials for investigating matrix interactions
| Reagent/Material | Function/Application | Experimental Context |
|---|---|---|
| Alginate Oligosaccharides (OligoG CF-5/20) | Matrix disruption agent; competes with native alginate for cationic bridging sites [60] | Studying matrix permeability, antibiotic potentiation (≥0.5% concentration) [60] |
| N-Acetyl Cysteine (NAC) | Biofilm disinfectant; kills embedded bacteria while preserving matrix structure for study [6] | Creating remnant matrices for recolonization studies (10 mg/mL concentration) [6] |
| POPC:POPG (70:30) Nanodiscs | Membrane mimic for structural studies of membrane-associated complexes [9] | Cryo-EM sample preparation for PelBC export complex [9] |
| Texas Red (TxRd) Conjugates | Fluorescent labeling for tracking molecular diffusion within biofilms [60] | Visualizing oligosaccharide penetration in established biofilms [60] |
| MSP1D1 Scaffold Protein | Forms lipid nanodiscs with controlled diameter (~8 nm) for membrane protein stabilization [9] | Reconstitution of membrane protein complexes for structural biology [9] |
Figure 2: Experimental workflow integrating key research reagents for investigating matrix interactions
The fortification of the P. aeruginosa biofilm matrix emerges from sophisticated synergy between its major components, governed by specific chemical interactions that include electrostatic binding, cationic bridging, and polyelectrolyte effects. The charge complementarity between cationic Pel and anionic eDNA and alginate creates a resilient network enhanced by divalent cation-mediated cross-linking [2] [60]. The functional consequence of these interactions is a matrix with tailored mechanical properties that resists antibiotic penetration and host immune clearance while maintaining metabolic flexibility [6] [69]. Current research tools—from high-resolution structural biology to functional microrheology—continue to reveal the complexity of these interactions, providing promising targets for therapeutic intervention such as alginate oligosaccharides that competitively disrupt critical cross-linking networks [60]. Future research focusing on the spatial organization of these interactions within intact biofilms and their dynamics during infection will further advance our ability to disrupt this protective matrix.
The biofilm matrix of Pseudomonas aeruginosa is a key determinant of its success as a pathogen, contributing significantly to chronic infections in cystic fibrosis (CF) airways, wounds, and implanted medical devices [16]. This extracellular matrix is primarily composed of three self-secreted exopolysaccharides (EPS)—alginate, Psl, and Pel—which collectively provide structural integrity, mediate adhesion, and confer formidable resistance to antimicrobials and host immune responses [16] [70]. While in vitro models have been instrumental in elucidating the fundamental biology of these EPS components, translating these findings to predict clinical isolate behavior in vivo presents a significant challenge in therapeutic development. This whitepaper examines the correlation between laboratory findings and clinical isolate behavior, providing a technical guide for researchers and drug development professionals working within the broader context of P. aeruginosa biofilm matrix research.
Exopolysaccharide Functions and Experimental Discrepancies
Distinct Functional Roles of Alginate, Psl, and Pel
The three major exopolysaccharides of P. aeruginosa perform complementary yet distinct functions in biofilm establishment and maintenance, though their relative importance varies considerably between strains and environmental conditions [17] [16].
Table 1: Key Functional Characteristics of P. aeruginosa Exopolysaccharides
| Exopolysaccharide | Chemical Composition | Primary Functions | Role in Antibiotic Resistance |
|---|---|---|---|
| Alginate | Linear copolymer of β-D-mannuronic and α-L-guluronic acids [66] | Predominant EPS in mucoid CF isolates; ROS scavenging; biofilm matrix scaffolding [66] [16] | Contributes to decreased susceptibility to antibiotics and host defenses [7] [16] |
| Psl | Neutral, branched pentasaccharide of D-mannose, D-glucose, and L-rhamnose [16] | Initial surface attachment; microcolony formation; structural integrity through cross-linking [7] [16] | Prevents complement deposition; inhibits neutrophil ROS production; sequesters antibiotics [16] |
| Pel | Cationic, partially de-N-acetylated polymer of α-1,4-N-acetylgalactosamine [9] [66] | Cell-cell interactions; pellicle formation at air-liquid interface; structural scaffold [7] [66] | Confers specific protection against aminoglycoside antibiotics [71] [3] |
Strain-Specific Variability in EPS Dependence
Significant functional divergence exists between laboratory strains and clinical isolates regarding their EPS utilization. Research demonstrates that P. aeruginosa can be categorized into at least four distinct classes based on their Pel and Psl functional and expression profiles [17]. For instance, the common laboratory strain PAO1 primarily utilizes Psl for biofilm structural integrity, while strain PA14 relies predominantly on Pel due to a natural deletion in its psl operon [17] [3]. This strain-specific variability extends to clinical isolates, with only approximately 30% of tested strains showing biofilm defects when pel is disrupted, whereas psl disruption consistently impairs initial surface attachment across diverse isolates [17].
In Vitro versus In Vivo Functional Correlations
The functional importance of specific EPS components observed in vitro does not always translate directly to in vivo settings. While Psl demonstrates critical importance for initial attachment in vitro across most strains [17], studies using mouse wound models revealed that neither deletion nor overproduction of Pel or Psl significantly affected bacterial load or wound closure rates [63]. However, the spatial organization of biofilm aggregates in wound tissue was markedly different when these EPS were absent, with smaller aggregates spaced further apart [63]. This suggests that EPS components may play a more subtle role in infection contexts where multiple adhesion mechanisms are available.
Table 2: Comparative EPS Functions in In Vitro vs. In Vivo Models
| Exopolysaccharide | Validated In Vitro Functions | Clinically Relevant In Vivo Observations |
|---|---|---|
| Psl | Essential for surface attachment in most strains; microcolony formation; structural scaffold in PAO1 [17] [3] | Presence in sputum aggregates associated with failed tobramycin eradication in CF [16]; affects aggregate size/distribution in wounds [63] |
| Pel | Structural scaffold in PA14; pellicle formation; aminoglycoside protection [3] | Contributes to biofilm cell density/compactness [7]; affects aggregate size in wounds; aminoglycoside tolerance in CF models [71] [63] |
| Alginate | ROS scavenging; antioxidant protection [66] | Associated with chronic CF infections; decreased susceptibility to antibiotics and host defenses [7] [16] |
Experimental Methodologies for EPS Characterization
Structural Analysis of EPS Secretion Complexes
Advanced structural biology techniques have been employed to characterize the molecular machinery responsible for EPS secretion. Recent cryo-EM studies of the Pel export complex (PelBC) have revealed its architecture at 2.5 Å resolution in lipid-based nanodiscs [9]. The experimental workflow involved:
- Membrane Protein Localization: Heterologous expression in E. coli C41(DE3) ΔompF ΔacrAB strain with verification of outer membrane localization via sucrose density gradient ultracentrifugation [9]
- Complex Extraction and Purification: Extraction with mild non-ionic detergent DDM followed by IMAC purification based on PelB's histidine tag [9]
- Membrane Mimicry: Reconstitution into POPC:POPG (70:30 molar ratio) nanodiscs using scaffold protein MSP1D1 to preserve native complex architecture [9]
- Structural Determination: Cryo-EM analysis of nanodisc-stabilized complexes with single-particle reconstruction [9]
This methodology revealed the 1:12 stoichiometry of the PelB:PelC complex and identified the gating mechanism for polysaccharide transport, providing insights critical for targeting EPS secretion therapeutically [9].
In Vitro Biofilm Phenotyping Assays
Standardized in vitro methods enable systematic evaluation of EPS contributions to biofilm development:
Microtiter Dish Biofilm Assay [3]:
- Grow mid-exponential phase cultures and dilute to OD650 0.1
- Aliquot 1 mL into 24-well polystyrene plates
- Incubate statically at 37°C for specified duration (typically 6-48 hours)
- Remove planktonic cells and stain adherent biomass with 0.1% crystal violet
- Quantify via solubilization with 30% acetic acid and measure A550
Flow Cell Reactor Biofilm Analysis [17] [3]:
- Grow biofilms in continuous flow chambers with dilute nutrient medium
- Monitor biofilm development in real-time using confocal laser scanning microscopy (CLSM)
- Analyze biofilm architecture, thickness, and biomass using image analysis software
- Assess spatial organization of different EPS components using fluorescent lectins or antibodies
In Vivo Infection Models
Multiple animal models have been developed to evaluate EPS function in clinically relevant contexts:
Mouse Wound Infection Model [63]:
- Create surgical excision wounds on mouse dorsum
- Infect with 1×10^6 CFU of P. aeruginosa wild-type or EPS mutants
- Monitor bacterial load in wound beds and spleens on days 2 and 7 post-infection
- Measure wound closure rates daily for 14 days
- Process tissue for histological analysis of biofilm aggregate distribution
Drosophila melanogaster Infection Model [71]:
- Infect flies via thorax puncture or natural feeding
- Monitor survival and bacterial persistence under antibiotic pressure
- Assess tolerance to clinically relevant antibiotics like tobramycin
Airway Epithelial Cell Colonization Model [71]:
- Culture wild-type (16HBEs) or CF (CFBEs) epithelial cells
- Infect with P. aeruginosa at specified MOI
- Assess bacterial persistence post-antibiotic treatment
- Measure cytotoxicity via LDH release assay
Signaling Pathways and Regulatory Networks in EPS Production
The production of Pel, Psl, and alginate is coordinately regulated through complex signaling networks that respond to environmental cues. Understanding these pathways is essential for predicting how clinical isolates may modulate EPS production in different infection contexts.
Environmental stressors prevalent in infection contexts significantly influence EPS production. Sublethal UVA exposure increases pelA and pslA gene expression, enhancing cell hydrophobicity and attachment capacity [66]. Nutrient availability also plays a crucial role; glycerol metabolism through the Glp regulon promotes Pel production and enhances tobramycin tolerance in CF-adapted isolates [71]. These regulatory interconnections create a sophisticated adaptation system that enables clinical isolates to modify their EPS profile according to specific host environments.
The Scientist's Toolkit: Essential Research Reagents and Models
Table 3: Key Research Reagents and Experimental Models for EPS Studies
| Category | Specific Reagents/Models | Research Applications | Technical Considerations |
|---|---|---|---|
| Bacterial Strains | PAO1 (Psl-dominant), PA14 (Pel-dominant), CF clinical isolates, EPS mutant panels (Δpel, Δpsl, Δalg) [17] [3] | Strain-specific EPS function; virulence studies; antibiotic tolerance mechanisms | Verify EPS production profile; monitor for compensatory mutations during culture [17] |
| Genetic Tools | araC-PBAD inducible systems [3], markerless deletion mutants [7], fluorescent reporter constructs | Controlled EPS expression; genetic screening; spatial localization in biofilms | Polar effects in operon deletions; epigenetic regulation of EPS genes |
| Biofilm Assay Systems | Static microtiter plates [3], flow cell reactors [17], pellicle formation assays [3] | Quantification of attachment and biofilm biomass; architectural analysis; air-liquid interface biofilms | Material surface properties affect attachment; nutrient composition influences EPS production |
| In Vivo Models | Mouse wound infection [63], Drosophila melanogaster [71], airway epithelial cell models [71] | Assessment of host-pathogen interactions; antibiotic efficacy; bacterial persistence | Host-specific factors influence EPS importance; model-dependent immune responses |
| Analytical Tools | Cryo-EM with nanodiscs [9], confocal microscopy with EPS-specific probes, lectin staining, RT-PCR for EPS gene expression | Structural biology of secretion complexes; spatial organization of EPS; gene regulation studies | Sample preparation critical for structural studies; probe specificity and accessibility |
Discussion and Future Perspectives
The correlation between in vitro findings and clinical isolate behavior remains complex due to strain-specific EPS utilization, environmental regulation of EPS production, and context-dependent functionality of matrix components. While in vitro models consistently demonstrate the importance of Psl for initial surface attachment and Pel for aminoglycoside protection [17] [3], in vivo observations reveal more nuanced roles where EPS components affect aggregate spatial distribution without necessarily altering overall bacterial burden [63].
Future research directions should prioritize:
- Development of more sophisticated in vitro models that better recapitulate host environments, including synthetic sputum media and co-culture systems
- Standardized comparative studies evaluating isogenic mutants across multiple in vitro and in vivo models
- Investigation of EPS functional redundancy and compensatory mechanisms in clinical isolates
- Exploration of EPS-targeting therapeutic adjuvants that can potentiate conventional antibiotics [16]
The continuing challenge in correlating laboratory findings with clinical isolate behavior underscores the necessity for multi-model validation approaches in therapeutic development targeting P. aeruginosa biofilms. As our understanding of EPS regulation and function improves, so too will our ability to predict how matrix-targeting strategies will perform across diverse clinical contexts.
The extracellular polysaccharides (EPS) alginate, Psl, and Pel are fundamental structural components of Pseudomonas aeruginosa biofilms, significantly influencing infection pathogenesis. This review synthesizes clinical and experimental evidence linking specific EPS profiles to infection outcomes in cystic fibrosis (CF) and chronic wound models. While alginate overproduction is a hallmark of chronic CF lung infections, its direct correlation with patient morbidity is complex. In contrast, Psl and Pel demonstrate critical, context-dependent roles in biofilm architecture, bacterial persistence, and antimicrobial tolerance. Through a systematic analysis of quantitative data from in vitro, in vivo, and clinical studies, this whitepaper establishes that EPS profiles are key determinants of infection trajectory, yet their impact is modulated by bacterial strain, infection site, and host environment. These insights are vital for guiding the development of targeted anti-biofilm therapeutics.
Pseudomonas aeruginosa is a formidable opportunistic pathogen whose resilience in chronic infections is largely attributable to its biofilm mode of growth. The biofilm matrix provides structural integrity and protects embedded cells from antibiotics and host immune defenses [2]. The three major exopolysaccharides—alginate, Psl, and Pel—are cornerstone components of this matrix. Within the context of a broader thesis on the role of these polymers in P. aeruginosa biofilm research, this review focuses specifically on elucidating their clinical correlations.
- Alginate: A high molecular weight, acetylated polymer of D-mannuronic and L-guluronic acid, alginate is classically associated with the mucoid phenotype of P. aeruginosa isolates from patients with CF [4]. Its overproduction was historically hypothesized to be a primary driver of poor clinical outcomes in CF.
- Psl: A neutral pentasaccharide composed of D-mannose, L-rhamnose, and D-glucose, Psl is crucial for surface attachment, the formation of microcolonies, and the maintenance of biofilm three-dimensional structure in non-mucoid strains [2] [4].
- Pel: A cationic polymer of partially de-N-acetylated N-acetylgalactosamine and N-acetylglucosamine, Pel facilitates cell-to-cell interactions and cross-links with extracellular DNA (eDNA), contributing to biofilm mechanical stability and tolerance to aminoglycoside antibiotics [2] [9].
Despite a deep understanding of their biochemical roles, translating EPS functions to predictable infection outcomes in humans has proven challenging. This review dissects the relationship between EPS profiles and clinical trajectories in two key infection models: the CF airway and the chronic wound.
EPS in Cystic Fibrosis Airway Infections
The CF lung is a primary site of chronic P. aeruginosa infection. The dehydrated, mucus-laden airway creates a niche where biofilm formation is heavily selected for, making EPS production a critical fitness factor.
Alginate: The Classic Virulence Determinant with Nuanced Impact
The conversion to a mucoid, alginate-overproducing phenotype is a key adaptive step in the progression from intermittent to chronic CF lung infection. Alginate is thought to confer a survival advantage by forming a protective, anionic hydrogel that shields bacteria from phagocytosis and opsonization [2] [4]. However, direct clinical correlation studies have yielded complex results. A comprehensive analysis of 108 BCC isolates from a Portuguese CF center found that while 80-90% of isolates produced the EPS cepacian, no clear correlation could be established between the in vitro ability to produce EPS and/or form biofilms and the persistence or virulence of the respiratory infections in different patients [72]. This suggests that other strain-dependent and host-specific factors are critical determinants of clinical outcomes.
Furthermore, studies on transmissible P. aeruginosa strains in CF patients have shown that infection with certain epidemic strains is not an independent risk factor for poorer post-lung transplantation outcomes, indicating that the inherent virulence of a strain, potentially linked to its EPS arsenal, must be considered within a broader pathophysiological context [73].
Psl and Pel: Contributors to Chronicity and Antibiotic Tolerance
While alginate dominates the narrative in CF, Psl and Pel are increasingly recognized for their roles in establishment and maintenance of infection. Psl is instrumental in the initial stages of infection, mediating attachment to the respiratory epithelium and abiotic surfaces [2]. It also contributes to antibiotic tolerance; Psl has been shown to limit the penetration of tobramycin and protect aggregates of P. aeruginosa in the sputum of children with CF, contributing to the failure of eradication therapy [2].
Pel, through its positive charge, binds negatively charged eDNA within the biofilm matrix. This interaction not only strengthens the biofilm structure but also protects the eDNA from degradation by DNases, potentially rendering nuclease-based therapies less effective [2]. This mechanism contributes significantly to aminoglycoside tolerance, a major therapeutic challenge in CF management.
Table 1: Correlation of EPS Type with Clinical and In Vitro Outcomes in CF
| EPS Type | Role in CF Biofilm | Impact on Antibiotic Tolerance | Correlation with Clinical Outcome |
|---|---|---|---|
| Alginate | Dominant in chronic/mucoid isolates; forms protective hydrogel matrix. | Confers broad-spectrum tolerance via impaired antibiotic penetration. | Associated with chronicity, but direct correlation with persistence/virulence is not clear-cut [72]. |
| Psl | Critical for initial adhesion & microcolony formation; maintains structure. | Sequesters tobramycin; prevents complement deposition, inhibiting phagocytosis [2]. | Linked to failure of tobramycin eradication therapy in early infection [2]. |
| Pel | Promotes cell-cell adhesion; cross-links eDNA for structural stability. | Confers specific tolerance to cationic aminoglycosides (e.g., tobramycin) [2]. | Contributes to bacterial persistence in the face of aminoglycoside treatment. |
EPS in Wound Model Infections
Chronic wounds, such as diabetic foot ulcers and venous leg ulcers, represent another clinical scenario where P. aeruginosa biofilms impede healing. In vivo wound models provide critical insights into how EPS composition influences infection pathogenesis outside the lung environment.
In Vivo Mouse Model Findings
A pivotal study using a murine wound infection model compared wild-type P. aeruginosa PAO1 (which produces Psl and Pel) with isogenic mutants lacking Pel (Δpel), Psl (Δpsl), or both (ΔpelΔpsl) [63]. The findings challenge simplistic assumptions about EPS and virulence:
- Infection Load and Healing Rate: The absence or overexpression of Pel and Psl did not significantly alter the bacterial load recovered from wounds or the rate of wound closure [63]. This indicates that, in this model, these EPS are not primary determinants of the overall bacterial burden or the gross impairment of healing.
- Spatial Organization of Biofilms: Despite no change in overall load, the spatial organization of bacteria in the wound tissue was profoundly affected. The ΔpelΔpsl double mutant formed significantly smaller bacterial aggregates that were spaced further apart compared to the wild-type strain [63].
- Antibiotic Survival: The ΔpelΔpsl double mutant demonstrated a significantly impaired ability to survive antibiotic treatment in vivo, highlighting the critical role of these EPS in mediating antimicrobial tolerance [63].
Insights from In Vitro Wound Slough Models
The development of an Artificial Wound Slough (AWS)-biofilm model has further elucidated the challenge of treating biofilm-associated wounds. This model, which involves growing a biofilm beneath a proteinaceous slough layer mimicking that found in chronic wounds, demonstrates that the physical barrier of slough and the embedded biofilm matrix severely limit the penetration of antimicrobial agents like silver [74]. This underscores the combined role of the host-derived slough and bacterial-derived EPS in creating a fortified, tolerant bacterial community.
Table 2: Impact of Pel and Psl on P. aeruginosa in a Murine Wound Model [63]
| Parameter Measured | Observation in Δpel, Δpsl, and ΔpelΔpsl mutants | Clinical Implication |
|---|---|---|
| Bacterial Load | No significant difference from wild-type PAO1. | Pel/Psl are not major virulence factors for establishing infection or growth in this model. |
| Wound Closure Rate | No significant difference from wild-type PAO1. | The presence of Pel/Psl does not directly dictate the macroscopic healing trajectory. |
| Aggregate Size & Distribution | Double mutant (ΔpelΔpsl) formed smaller, more dispersed aggregates. | EPS are key determinants of biofilm architecture and community cohesion in vivo. |
| Survival after Antibiotic Treatment | Double mutant (ΔpelΔpsl) showed significantly reduced survival. | Pel and Psl are critical for antibiotic tolerance and bacterial persistence during treatment. |
Experimental Protocols for EPS and Biofilm Analysis
To standardize research in this field, detailed methodologies for key experiments are provided below.
Static Biofilm Formation Assay (Microtiter Plate)
This is a foundational protocol for quantifying biofilm formation in vitro [72].
- Inoculum Preparation: Grow bacteria overnight in a suitable liquid medium (e.g., Lennox broth). Dilute the culture to a standardized optical density at 640 nm (OD₆₄₀) of 0.5.
- Inoculation: Transfer 20 µL of the standardized cell suspension into the wells of a 96-well polystyrene microtiter plate containing 180 µL of fresh medium. Include wells with sterile medium as negative controls.
- Incubation: Incubate the plate without agitation at a desired temperature (e.g., 30°C or 37°C) for a defined period (e.g., 24 or 48 hours).
- Biofilm Quantification (Crystal Violet Staining): a. Carefully remove the planktonic culture and rinse the wells three times with 200 µL of water to remove non-adherent cells. b. Stain the adherent biofilms with 200 µL of a 1% (w/v) crystal violet solution for 15 minutes at room temperature. c. Rinse the wells three times with 200 µL of water to remove unbound dye. d. Solubilize the crystal violet bound to the biofilm in 200 µL of 95% ethanol. e. Quantify the biofilm by measuring the absorbance of the solubilized dye at 590 nm (A₅₉₀) using a microplate reader.
In Vivo Murine Wound Infection Model
This protocol outlines the procedure for assessing the role of EPS in a live wound environment [63].
- Animal Preparation: Anesthetize mice and create full-thickness excision wounds on the dorsal surface.
- Infection: Inoculate the wound bed with a defined inoculum (e.g., 10⁵ CFU) of the P. aeruginosa strain of interest (e.g., wild-type, Δpel, Δpsl, ΔpelΔpsl).
- Monitoring:
- Bacterial Load: At predetermined time points (e.g., days 2 and 7), euthanize a cohort of animals. Excise the entire wound and homogenize the tissue. Plate serial dilutions of the homogenate to determine the bacterial load (CFU/wound).
- Wound Closure: Photograph the wounds daily and measure the wound area using image analysis software to calculate the rate of wound closure over time (e.g., 14 days).
- Histology and Imaging: For spatial analysis of biofilms, excise wound tissue, process for histology (e.g., fixation, sectioning), and stain with fluorescent probes (e.g., FISH, antibodies) for examination by confocal or fluorescence microscopy.
Artificial Wound Slough (AWS)-Biofilm Model
This in vitro model is designed to mimic the slough-covered biofilms in chronic wounds [74].
- AWS Preparation: Prepare a homogenized suspension of artificial slough containing 65% collagen, 10% elastin, and 25% fibrinogen. Clot the solution by mixing with thrombin (6.25 U/mL) in a 1:1 (v/v) ratio.
- Biofilm Establishment: Inoculate wells of a 48-well plate with P. aeruginosa (e.g., 400 µL of 10⁶ CFU/mL) and incubate for 24 hours at 37°C to form a base biofilm.
- Slough Overlay: Aspirate the spent medium and gently wash the biofilm. Add the AWS solution on top of the established biofilm to create layers of defined thickness (e.g., 0.5 mm, 1.0 mm, 2.0 mm). Incubate the plate at 37°C with gentle shaking to set.
- Antimicrobial Efficacy Testing: Apply the antimicrobial treatment (e.g., silver solution) on top of the AWS layer. After incubation, disrupt the AWS-biofilm composite, vortex, and perform serial dilution and plating to quantify viable bacteria beneath the slough layer.
Visualization of EPS Biosynthesis and Experimental Workflows
EPS Biosynthesis and Secretion Pathway in P. aeruginosa
Workflow for In Vivo Wound Infection Analysis
The Scientist's Toolkit: Key Research Reagents and Models
Table 3: Essential Research Tools for Investigating P. aeruginosa EPS
| Tool / Reagent | Function / Application | Example Use |
|---|---|---|
| Isogenic Mutant Strains (e.g., ΔpelA, ΔpslBCD, Δalg8, ΔwspF) | To define the specific function of a single EPS by comparing mutant phenotypes to wild-type. ΔwspF hyperproduces Pel and Psl [63]. | Phenotypic characterization of biofilm formation, architecture, and antibiotic tolerance [63] [8]. |
| Crystal Violet Staining | A high-throughput method for quantifying total adhered biofilm biomass in microtiter plates [72]. | Initial screening of biofilm-deficient or -enhanced mutants [72] [8]. |
| Confocal Laser Scanning Microscopy (CLSM) | Enables 3D, non-destructive imaging of hydrated biofilm architecture, thickness, and biovolume. | Visualizing the fabric-like Psl matrix [4] and quantifying structural changes in mutant biofilms [8]. |
| Artificial Wound Slough (AWS) Model | An in vitro model that mimics the proteinaceous slough covering biofilms in chronic wounds [74]. | Testing antimicrobial penetration and efficacy against biofilms protected by a physical barrier [74]. |
| Quartz Crystal Microbalance with Dissipation (QCM-D) | Label-free, real-time monitoring of biofilm mass, thickness, and viscoelastic properties. | Demonstrating that Psl is the primary driver of biofilm elasticity and maturation [8]. |
| Murine Wound Infection Model | An in vivo system for studying the role of EPS in pathogenesis, persistence, and host response. | Revealing that Pel/Psl dictate aggregate structure and antibiotic survival, but not overall bacterial load [63]. |
The correlation between EPS profiles and infection outcomes is unequivocal yet context-dependent. In CF, alginate production is a marker of chronic adaptation, but Psl and Pel are critical for the establishment and antibiotic tolerance of the biofilm. In wound models, Psl and Pel are non-essential for initiating infection but are indispensable for structuring robust biofilms and surviving antimicrobial treatment. The hierarchical importance of Psl in biofilm mechanics, as revealed by multi-omics approaches [8], positions it as a prime therapeutic target. Future anti-biofilm strategies must move beyond broad-spectrum approaches and instead target the specific EPS pathways that drive persistence in a given clinical context. The experimental tools and data synthesized in this review provide a foundation for such targeted therapeutic development.
Conclusion
The exopolysaccharides alginate, Psl, and Pel are not merely inert structural components but are dynamic, multifunctional guardians of the P. aeruginosa biofilm community. Their roles are distinct yet deeply intertwined, providing a robust and redundant matrix system that confers formidable resistance to antimicrobials and host defenses. While alginate is a classic virulence factor in chronic infections, Psl and Pel are now recognized as equally critical for biofilm integrity and tolerance. The future of combating P. aeruginosa infections lies in moving beyond conventional antibiotics towards combinatorial strategies that simultaneously disrupt these EPS networks. Promising avenues include the clinical development of EPS-degrading enzymes as potent antibiotic adjuvants and the manipulation of regulatory pathways like c-di-GMP signaling. A profound, integrated understanding of this polysaccharide matrix is the key to unlocking novel, effective therapies against one of the most resilient bacterial pathogens.
References
- 1. Role of polysaccharides in Pseudomonas aeruginosa ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2176169/]
- 2. How Three Self-Secreted Biofilm Exopolysaccharides of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10218432/]
- 3. The Pel Polysaccharide Can Serve a Structural and Protective ... [https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1001264]
- 4. Biosynthesis of the Pseudomonas aeruginosa Extracellular ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2011.00167/full]
- 5. The Pel polysaccharide is predominantly composed of a ... [https://www.nature.com/articles/s42003-022-03453-2]
- 6. Alginate exopolymer significantly modulates the ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12149293/]
- 7. Role of Exopolysaccharides in Pseudomonas aeruginosa ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3147449/]
- 8. New Insights into the Mediation of Biofilm Formation by ... [https://www.mdpi.com/1422-0067/26/8/3780]
- 9. Assembly and the gating mechanism of the Pel ... [https://www.nature.com/articles/s41467-025-60605-8]
- 10. Biosynthesis of the Pseudomonas aeruginosa Extracellular ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3159412/]
- 11. Insights into the Assembly of the Alginate Biosynthesis ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3685254/]
- 12. Structural and functional analysis of Pseudomonas ... [https://www.sciencedirect.com/science/article/pii/S0021925825002819]
- 13. Alginate exopolymer significantly modulates the ... [https://www.nature.com/articles/s41522-025-00718-6]
- 14. New Insights into the Mediation of Biofilm Formation by Three ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12027665/]
- 15. Immune system dynamics in response to Pseudomonas ... [https://www.nature.com/articles/s41522-025-00738-2]
- 16. How Three Self-Secreted Biofilm Exopolysaccharides of ... [https://www.mdpi.com/1422-0067/24/10/8709]
- 17. The Pel and Psl polysaccharides provide Pseudomonas ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3840794/]
- 18. The roles of Pseudomonas aeruginosa extracellular ... [https://www.sciencedirect.com/science/article/abs/pii/S0376738814003457]
- 19. Psl Produced by Mucoid Pseudomonas aeruginosa ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5478896/]
- 20. Cellulophaga algicola alginate lyase and Pseudomonas ... [https://www.sciencedirect.com/science/article/pii/S2590207525000139]
- 21. New research uncovers how bad bacteria know where to ... [https://cnsi.ucla.edu/october-28-2025-new-research-uncovers-how-bad-bacteria-know-where-to-cluster-and-cause-infection/]
- 22. Specific Disruption of Established P. aeruginosa Biofilms ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7063831/]
- 23. Two unrelated Pseudomonas aeruginosa phages require ... [https://www.nature.com/articles/s41522-025-00841-4]
- 24. Directed evolution of phages in biofilms enhances ... [https://www.nature.com/articles/s41467-025-65014-5]
- 25. Pseudomonas aeruginosa Biofilms - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC7698413/]
- 26. Biofilm Matrix and Its Regulation in Pseudomonas ... [https://www.mdpi.com/1422-0067/14/10/20983]
- 27. Pseudomonas biofilm matrix composition and niche biology [https://pmc.ncbi.nlm.nih.gov/articles/PMC4409827/]
- 28. Assembly and Development of the Pseudomonas aeruginosa ... [https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1000354]
- 29. Biofilms and Cyclic di-GMP (c-di-GMP) Signaling [https://pmc.ncbi.nlm.nih.gov/articles/PMC4933438/]
- 30. Regulation of Biofilm Exopolysaccharide Production by ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2021.730980/full]
- 31. Cyclic-di-GMP Regulates Extracellular Polysaccharide ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2446529/]
- 32. A cyclic-di-GMP receptor required for bacterial ... - PubMed [https://pubmed.ncbi.nlm.nih.gov/17824927/]
- 33. Pseudomonas aeruginosa Biofilm Formation and Its Control [https://www.mdpi.com/2673-8449/1/3/19]
- 34. A novel multiplex fluorescent-labeling method for the ... [https://pubmed.ncbi.nlm.nih.gov/38785429/]
- 35. Imaging of bacterial multicellular behaviour in biofilms ... [https://www.nature.com/articles/srep25889]
- 36. The Pel and Psl polysaccharides provide Pseudomonas aeruginosa structural redundancy within the biofilm matrix - PubMed [https://pubmed.ncbi.nlm.nih.gov/22176658/]
- 37. Microtiter Dish Biofilm Formation Assay - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3182663/]
- 38. Interlaboratory study for the evaluation of three microtiter ... [https://www.nature.com/articles/s41598-021-93115-w]
- 39. Pseudomonas aeruginosa: Infections, Animal Modeling, and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9818774/]
- 40. The landscape of biofilm models for phage therapy [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2025.1553979/full]
- 41. Strategy to combat biofilms: a focus on biofilm dispersal ... [https://www.nature.com/articles/s41522-023-00427-y]
- 42. Role of exopolysaccharides in Pseudomonas ... aeruginosa biofilm [https://pubmed.ncbi.nlm.nih.gov/21666010/]
- 43. Extracellular Polysaccharides in Pseudomonas aeruginosa [https://digital.lib.washington.edu/researchworks/items/f541a8b4-1fe9-4997-9cc0-feb7046bbcda]
- 44. The pel polysaccharide can serve a structural and protective role in the... [https://pubmed.ncbi.nlm.nih.gov/21298031/]
- 45. The Pel and Psl polysaccharides provide Pseudomonas ... aeruginosa [https://researchportal.bath.ac.uk/en/publications/the-pel-and-psl-polysaccharides-provide-pseudomonas-aeruginosa-st]
- 46. (PDF) The Pel and Psl polysaccharides provide Pseudomonas ... [https://www.academia.edu/79891925/The_Pel_and_Psl_polysaccharides_provide_Pseudomonas_aeruginosa_structural_redundancy_within_the_biofilm_matrix]
- 47. Biofilm inhibitory effect of alginate lyases on mucoid P. ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8165544/]
- 48. minimally invasive therapy for biofilm penetration and ... [https://www.sciencedirect.com/science/article/pii/S2211383525001741]
- 49. The biofilm matrix | Nature Reviews Microbiology [https://www.nature.com/articles/nrmicro2415?error=cookies_not_supported]
- 50. Targeted disruption of the extracellular polymeric network ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6026129/]
- 51. Calcium Chelation by Alginate Activates the Type III... | PLOS One [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0046826]
- 52. Repurposing antimicrobials with ultrasound-triggered ... [https://www.nature.com/articles/s44259-025-00086-3]
- 53. Contribution of alginate and levan production to biofilm by... formation [https://pubmed.ncbi.nlm.nih.gov/17005972/]
- 54. Biofilm, resistance, and quorum sensing: The triple threat in ... [https://www.sciencedirect.com/science/article/pii/S2950194625003462]
- 55. Antibiotic potentiators as a promising strategy for ... [https://www.nature.com/articles/s44259-025-00112-4]
- 56. Mechanism, Efficacy, and Safety of Natural Antibiotics [https://pubmed.ncbi.nlm.nih.gov/41148673/]
- 57. The Pel Polysaccharide Can Serve a Structural and Protective ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3029257/]
- 58. Characterization of different alginate lyases for dissolving ... [https://www.nature.com/articles/s41598-020-66293-2]
- 59. Prevalence of algD, pslD, and pelF Genes Involved in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12520592/]
- 60. Targeted disruption of the extracellular polymeric network ... [https://www.nature.com/articles/s41522-018-0056-3]
- 61. Building a better biofilm - Formation of in vivo-like ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7762787/]
- 62. Antimicrobial resistance of Pseudomonas aeruginosa [https://pmc.ncbi.nlm.nih.gov/articles/PMC11026690/]
- 63. Contribution of Pseudomonas aeruginosa ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9021892/]
- 64. The role of exopolysaccharides Psl and Pel in resistance ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11448232/]
- 65. Polysaccharides' Structures and Functions in Biofilm ... [https://www.mdpi.com/1422-0067/24/4/4030]
- 66. Role of Pel and Psl polysaccharides in the response ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10197878/]
- 67. The pattern of antibiotic resistance and distribution ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11569483/]
- 68. Pseudomonas aeruginosa polysaccharide Psl supports ... [https://www.nature.com/articles/s41396-022-01221-y]
- 69. Pseudomonas aeruginosa Biofilms [https://www.mdpi.com/1422-0067/21/22/8671]
- 70. Biofilms: structure, resistance mechanism, emerging control ... [https://pubs.rsc.org/en/content/articlehtml/2025/pm/d5pm00094g]
- 71. Activation of the Pseudomonas aeruginosa glycerol ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12519775/]
- 72. Studies on the Involvement of the Exopolysaccharide ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC446245/]
- 73. Epidemic Pseudomonas aeruginosa infection in patients ... [https://www.sciencedirect.com/science/article/pii/S1569199315002623]
- 74. An In Vitro Artificial Wound Slough–Biofilm Model ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11596274/]