Polysaccharide Intercellular Adhesin (PIA): Structure, Function, and Therapeutic Targeting in Staphylococcal Biofilms
This article comprehensively reviews the polysaccharide intercellular adhesin (PIA), a key exopolysaccharide component of the Staphylococcus epidermidis and Staphylococcus aureus biofilm matrix.
Polysaccharide Intercellular Adhesin (PIA): Structure, Function, and Therapeutic Targeting in Staphylococcal Biofilms
Abstract
This article comprehensively reviews the polysaccharide intercellular adhesin (PIA), a key exopolysaccharide component of the Staphylococcus epidermidis and Staphylococcus aureus biofilm matrix. We explore PIA's structural characterization as a poly-β(1-6)-N-acetylglucosamine (PNAG), its biosynthesis via the icaADBC locus, and its dual role in mediating biofilm accumulation and immune evasion. For researchers and drug development professionals, we detail methodological approaches for PIA investigation, analyze PIA-independent biofilm mechanisms, and evaluate PIA's validation as a therapeutic target through vaccine development and anti-biofilm strategies. The synthesis of current evidence positions PIA at the forefront of innovative approaches to combat biomaterial-associated infections.
Unraveling PIA: Structural Identity and Biosynthetic Machinery in Staphylococci
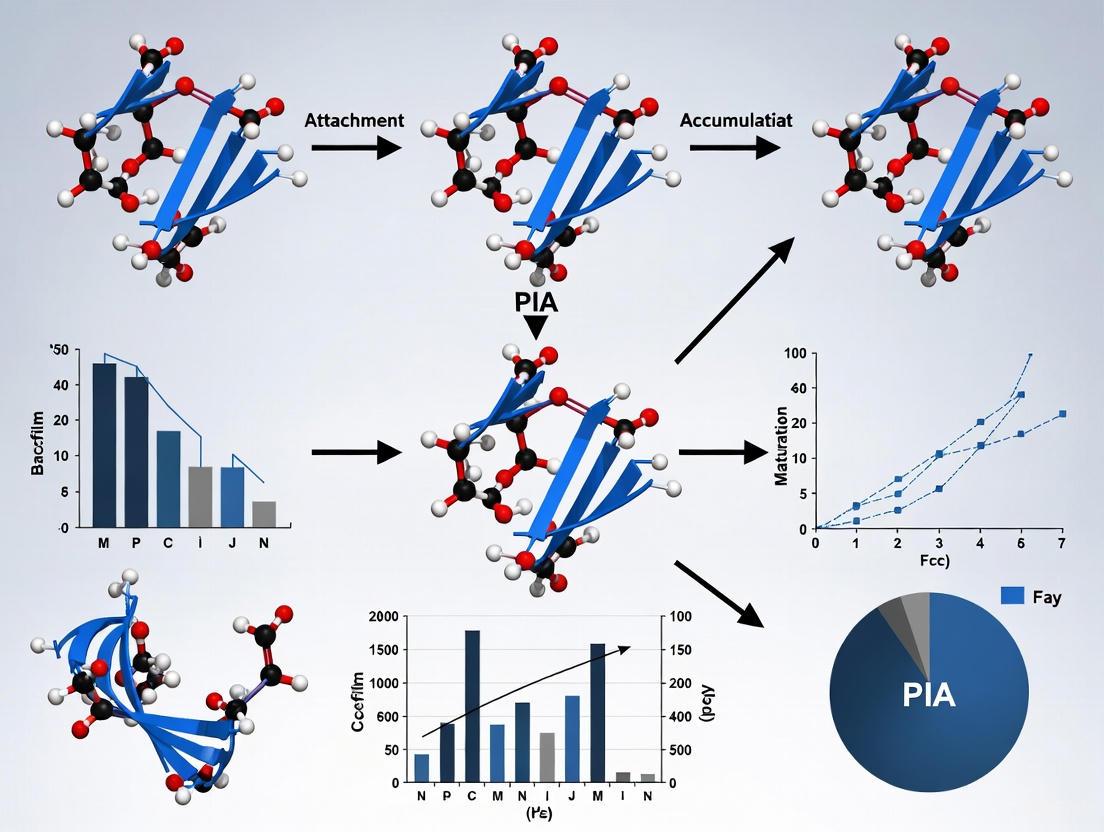
Polysaccharide intercellular adhesin (PIA) is a crucial component of the extracellular matrix in staphylococcal biofilms, serving as a primary virulence factor in implant-associated infections. Staphylococcus aureus and Staphylococcus epidermidis, the leading etiologic agents of medical device-related infections, rely on PIA for the structural integrity of their biofilms, which significantly increases their resistance to antibiotics and host immune defenses [1] [2]. This exopolysaccharide was initially identified in S. epidermidis and has since been recognized as a fundamental factor in the pathogenesis of chronic infections [3]. The chemical identity of PIA was definitively established in 1996 as a linear, partially deacetylated polymer of β-1-6-linked N-acetylglucosamine (GlcNAc), forming a poly-β(1-6)-N-acetylglucosamine (PNAG) structure [3]. This architectural foundation enables PIA to facilitate cell-to-cell adhesion, promote biofilm maturation, and provide a protective barrier against external threats, establishing it as a critical research focus in developing anti-biofilm therapeutic strategies.
Chemical Structure and Molecular Characteristics
The PIA polymer consists of a linear chain of D-glucosamine residues connected exclusively by β-1,6-glycosidic linkages. The fundamental monomeric unit is N-acetylglucosamine (GlcNAc), an amide derivative of glucose where the C-2 hydroxyl group is replaced by an acetamido function [4]. In its nascent form, immediately after biosynthesis, the polymer is a fully acetylated poly-β-1,6-N-acetylglucosamine (PNAG). However, the mature, functional exopolysaccharide undergoes essential enzymatic modification: approximately 15-20% of the N-acetylglucosamine residues are deacetylated by the enzyme IcaB, generating positively charged amino groups (GlcNH₃⁺) along the polymer backbone [3] [5]. This partial deacetylation is not random but appears to occur in distinct regions, creating a mosaic of highly acetylated and highly deacetylated domains within the biofilm architecture [6].
Additional structural modifications have been reported, though their prevalence and functional significance can vary. O-succinylation of a portion (approximately 10%) of the sugar residues has been documented in PIA from both S. epidermidis and S. aureus [3]. Early reports suggested N-succinylation, but these were later identified as analytical artifacts [3]. The polymer size displays considerable heterogeneity, with molecular weight estimates ranging from ~30 kDa (approximately 130 residues) to over 460 kDa, depending on the bacterial strain, growth conditions, and isolation methods [3]. The resulting cationic character of the partially deacetylated PIA, due to the protonated amino groups, is critical for its adherence to the anionic bacterial cell surface and for its role in intercellular adhesion [6] [5].
Table 1: Molecular Characteristics of Polysaccharide Intercellular Adhesin (PIA)
| Characteristic | Description | Functional Significance |
|---|---|---|
| Chemical Identity | Poly-β(1-6)-N-acetylglucosamine (PNAG) | Defines the polymer backbone structure [3] |
| Glycosidic Linkage | β-1,6 | Determines linear chain conformation and physical properties [3] |
| Native Monomer | N-acetylglucosamine (GlcNAc) | Building block for polymerization [4] |
| Key Modification | Partial (~15-20%) enzymatic de-N-acetylation | Generates positive charge for surface attachment and biofilm integrity [3] [5] |
| Reported Size Range | ~30 kDa to >460 kDa | Heterogeneity depends on species, strain, and growth conditions [3] |
| Net Charge | Cationic (after deacetylation) | Essential for adhesion to anionic cell surfaces [6] |
Biosynthetic Machinery and Regulatory Elements
The synthesis, export, and modification of PIA are governed by the icaADBC operon. Each gene in this locus encodes a protein with a distinct and essential function, working in concert to produce the mature exopolysaccharide.
- IcaA: Functions as the core N-acetylglucosaminyltransferase. It utilizes the cytoplasmic substrate UDP-N-acetylglucosamine to catalyze the formation of β-1,6-glycosidic bonds, elongating the PNAG chain. IcaA is an integral membrane protein with a large cytosolic glycosyltransferase domain [7] [5].
- IcaD: A small integral membrane protein that substantially increases the catalytic efficiency of IcaA when co-expressed. While IcaA alone exhibits low transferase activity, the IcaA-IcaD complex produces PNAG oligomers up to 20 residues long in vitro, indicating that IcaD is crucial for optimal polymer synthesis [7].
- IcaC: An integral membrane protein predicted to contain multiple transmembrane helices. Initially thought to be an exporter for long-chain PNAG, recent bioinformatic analyses suggest it may belong to an acetyltransferase family and could be involved in the O-succinylation of the polysaccharide during its biosynthesis [5].
- IcaB: A secreted extracellular deacetylase belonging to the family 4 carbohydrate esterases (CE4s). IcaB is responsible for the partial de-N-acetylation of the exported, fully acetylated PNAG polymer. This enzymatic activity is a critical prerequisite for the polymer's retention on the bacterial cell surface and for the development of a robust biofilm [6] [5]. Strains with icaB knocked out produce fully acetylated PNAG that is not retained on the surface, fail to form biofilms, and are avirulent in infection models [5].
The expression of the icaADBC operon is tightly regulated by environmental conditions and various regulatory systems, including quorum sensing, which can negatively regulate biofilm formation to favor the dispersal phase for infection dissemination [1] [2].
Distribution Across Bacterial Species and Functional Role
PIA is not exclusive to staphylococci; it is a conserved exopolysaccharide produced by a wide range of Gram-positive and Gram-negative bacterial pathogens. In Escherichia coli and other Gram-negative bacteria, the homologous operon is designated pgaABCD, with PgaB serving the deacetylase function analogous to IcaB [8] [3]. However, a key structural difference exists: the Gram-negative PgaB requires a C-terminal domain for efficient deacetylation, whereas the staphylococcal IcaB does not, a distinction with important implications for inhibitor design [5]. Beyond E. coli, functional homologs have been biochemically confirmed in significant human pathogens such as Acinetobacter baumannii, Klebsiella pneumoniae, Yersinia pestis, Bordetella spp., and members of the Burkholderia cepacia complex [8] [3] [6].
In all these species, PIA plays a consistent role as a key biofilm matrix component. Its production is strongly linked to virulence, contributing to immune evasion by providing resistance to antimicrobial peptides and protecting against phagocyte ingestion and killing [3]. The polymer's positive charge facilitates adhesion to surfaces, and its physical structure creates a diffusion barrier, collectively enhancing bacterial persistence during chronic infections.
Table 2: Distribution of PIA/PNAG in Pathogenic Bacteria
| Bacterial Species | Genetic Locus | Confirmed Role in Biofilm | Key References |
|---|---|---|---|
| Staphylococcus epidermidis | icaADBC | Major biofilm matrix component; definitive virulence factor [3] [2] | [1] [3] |
| Staphylococcus aureus | icaADBC | Key component in many strains; contributes to immune evasion [3] [6] | [1] [6] |
| Escherichia coli | pgaABCD | Promotes biofilm formation and structure [8] [3] | [8] [3] |
| Burkholderia cepacia complex | pgaBCD (homolog) | Required for thick, mature biofilm architecture [8] | [8] |
| Acinetobacter baumannii | Homologs identified | Biofilm component and virulence factor [3] [6] | [3] [6] |
| Yersinia pestis | hmsHFRS | Critical for biofilm formation block of flea proventriculus [3] | [3] |
Experimental Analysis and Methodological Approaches
Immunoblot Analysis for PIA Detection
A standard method for detecting PIA involves immunoblot analysis using specific monoclonal antibodies. The polysaccharide is first extracted from bacterial cells harvested from biofilms. Cells are resuspended in 0.5 M EDTA (pH 8.0) and incubated at 100°C for 5 minutes followed by 85°C for 30 minutes [8]. The clarified supernatant is dialyzed extensively against deionized water and then against 50 mM Tris-HCl (pH 8.0) containing 20 mM MgCl₂. The crude extract is treated with a series of enzymes to remove contaminants: α-amylase (100 µg/mL) to degrade starch, lysozyme (500 µg/mL), DNase I (250 µg/mL), and RNase A (100 µg/mL) at 37°C for 2 hours, followed by digestion with proteinase K (2 mg/mL) for 16 hours at 55°C in the presence of 1 mM CaCl₂ and 0.5% SDS [8]. After enzyme inactivation at 85°C for 1 hour and further dialysis, the polysaccharide preparation is lyophilized. Samples are spotted onto a PVDF membrane, blocked, and probed with a primary monoclonal antibody (e.g., murine IgM against E. coli PGA at 1:2,000 dilution). Binding is detected using a horseradish peroxidase-conjugated secondary antibody (e.g., goat anti-mouse IgM at 1:10,000) and a chromogenic substrate like 3,3',5,5'-tetramethylbenzidine (TMB) [8].
Functional Biofilm Disruption Assays
The specific role of PIA in biofilms can be functionally assessed using the enzyme Dispersin B, a PNAG-specific hydrolase. To evaluate its inhibitory effect on biofilm formation, Dispersin B (with a specific activity of ~103 units/mg protein) is added to the growth medium at the initiation of bacterial culture [8]. To assess dispersal of pre-formed biofilms, the enzyme is added to mature biofilms and incubated for a defined period. Biofilm mass and architecture are then quantified and visualized using methods like crystal violet staining and confocal scanning laser microscopy (CSLM) [8]. Genetic confirmation involves constructing isogenic mutant strains, such as a ΔpgaBC mutant in B. multivorans or a ΔicaB mutant in staphylococci, which are expected to produce no PIA or a fully acetylated, non-adherent polymer, respectively, resulting in thin, poorly developed biofilms as seen by CSLM [8] [5].
Research Reagents and Therapeutic Applications
The study of PIA and the development of anti-biofilm strategies rely on a defined set of research reagents and tools. Key among these are specific enzymes and antibodies that enable the manipulation, detection, and functional characterization of this critical polysaccharide.
Table 3: Essential Research Reagents for PIA/PNAG Studies
| Reagent | Type | Specific Function in Research | Example Application |
|---|---|---|---|
| Dispersin B | Glycoside Hydrolase Enzyme | Hydrolyzes β-1,6-glycosidic linkages in PNAG [8] | Biofilm disruption assays; confirm PIA function [8] |
| Anti-PNAG mAb (e.g., F598) | Monoclonal Antibody | Binds to acetylated epitopes on native PIA; opsonic killing [6] | Immunoblot detection, microscopy, therapeutic studies [8] [6] |
| Anti-dPNAG mAb (e.g., TG10) | Monoclonal Antibody | Binds selectively to highly deacetylated PNAG epitopes [6] | Mapping deacetylated regions in biofilms; combination therapy [6] |
| IcaB/PgaB | Deacetylase Enzyme | Catalyzes de-N-acetylation of PNAG in vitro [5] | Biochemical studies of deacetylation mechanism and kinetics [5] |
| Synthetic dPNAG Glycans | Defined Oligosaccharide Antigens | Glycans with controlled chain length and acetylation [9] | Vaccine development; antibody response profiling via microarrays [9] |
The conserved nature and critical role of PIA in virulence have made it a prominent target for novel therapeutic and prophylactic approaches. Two main strategies have emerged: immunotherapy using monoclonal antibodies and active vaccination. The most advanced monoclonal antibody, F598, which binds both acetylated and deacetylated PNAG, has progressed to Phase II clinical trials as a broad-spectrum anti-biofilm agent [6]. Recent research indicates that a combination of antibodies, such as F598 (binding acetylated epitopes) and TG10 (binding deacetylated epitopes), can synergize to provide superior coverage and protection in animal models by targeting the distinct architectural regions of PIA within biofilms [6]. For vaccines, chemically or synthetically produced deacetylated PNAG (dPNAG) glycans conjugated to carrier proteins like CRM197 (a diphtheria toxin mutant) have shown great promise. These conjugate vaccines are designed to elicit a potent, opsonic antibody response that promotes bacterial clearance [9]. Systematic optimization has identified glycans with a degree of polymerization of 8 or 12 and approximately 40-45% acetylation as particularly effective immunogens, capable of inducing broad protection against multiple bacterial pathogens in preclinical models [9].
The icaADBC operon is a conserved genetic locus that serves as the fundamental blueprint for the synthesis of Polysaccharide Intercellular Adhesin (PIA), a critical virulence determinant in staphylococcal biofilms. Biofilm formation is a key pathogenicity strategy for bacteria like Staphylococcus aureus and Staphylococcus epidermidis, allowing them to adhere to medical devices and host tissues, and to resist antibiotic treatments and host immune responses [10] [11]. The extracellular matrix of these biofilms provides structural integrity and protection, with PIA being a major exopolysaccharide component [12]. This operon encodes the enzymatic machinery for the production of a linear β-1,6-linked N-acetylglucosamine polymer, which is essential for the cell-to-cell adhesion that characterizes biofilm accumulation [13] [14]. Understanding the function and regulation of this operon is paramount for developing novel anti-biofilm strategies aimed at treating persistent device-related infections.
Genetic Architecture and Molecular Functions of the icaADBC Operon
The icaADBC operon comprises four core genes—icaA, icaD, icaB, and icaC—which work in concert to synthesize and modify PIA. The coordinated activity of these gene products results in a functional polysaccharide that facilitates robust biofilm formation.
- icaA: Encodes for N-acetylglucosaminyl transferase, the primary transmembrane enzyme that catalyzes the synthesis of the oligomer chain of Poly-N-acetylglucosamine (PNAG) [15] [11]. Its enzymatic activity is the cornerstone of PIA production.
- icaD: Co-transcribed with icaA, its gene product acts as a chaperone protein that stabilizes IcaA and significantly enhances its enzymatic activity and specificity, leading to the production of longer, more functional polymers [15] [11].
- icaC: A transmembrane protein believed to be involved in the translocation and external elongation of the nascent PIA polymer across the bacterial cell membrane to the cell surface [15] [11].
- icaB: Encodes a deacetylase enzyme responsible for the critical maturation step of PIA. It partially deacetylates the polymer, imparting a net positive charge that is essential for its adherence to the negatively charged bacterial cell surface and for mediating intercellular adhesion [15] [11].
The table below summarizes the core components and their precise molecular functions.
Table 1: Core Genes of the icaADBC Operon and Their Functions
| Gene | Encoded Protein Function | Role in PIA Biosynthesis |
|---|---|---|
| icaA | N-acetylglucosaminyl transferase | Catalyzes polymerization of N-acetylglucosamine residues [15] [11] |
| icaD | Chaperone protein | Stabilizes IcaA, essential for full enzymatic activity and long-chain polymer synthesis [15] [11] |
| icaC | Transmembrane transporter | Facilitates PIA export and elongation on the cell surface [15] [11] |
| icaB | Deacetylase | Partially deacetylates PIA, conferring a positive charge for adhesion [15] [11] |
Diagram 1: PIA Biosynthesis Pathway. This diagram illustrates the coordinated roles of the icaADBC gene products in synthesizing, exporting, and modifying PIA to form the biofilm matrix.
Regulatory Mechanisms and Environmental Influence
The expression of the icaADBC operon is not constitutive but is under sophisticated control by a network of regulatory systems and environmental stimuli, which adds a layer of complexity to its role in virulence.
Transcriptional Regulation
The operon is repressed by IcaR, a transcriptional repressor encoded by a gene located upstream of the operon. Mutations in the icaR gene can lead to constitutive overexpression of the operon, resulting in a mucoid phenotype with excessive PIA production, as observed in isolates from cystic fibrosis patients [16]. Other global regulatory systems, including the accessory gene regulator (agr), staphylococcal accessory regulator (SarA), and the SaeRS two-component system, also intricately modulate ica expression [17] [11]. For instance, active agr typically downregulates biofilm formation, promoting a planktonic lifestyle [17].
Phase Variation via IS256
The IS256 insertion sequence plays a critical role in phase variation, a reversible on/off switching of the operon's activity. IS256 can insert into the icaA or icaC genes, disrupting their function and abolishing PIA production [13] [14]. This phase variation is considered a pathogenicity adaptation, allowing subpopulations to evade host defenses. Clinically, the presence of IS256 is strongly associated with therapeutic failure in device-related infections caused by coagulase-negative staphylococci [13].
Environmental Triggers
Environmental conditions significantly influence ica operon expression. Key inducing factors include:
- Anaerobic conditions and high glucose concentrations [10] [17].
- High salt concentrations, sub-inhibitory antibiotic stress, and the presence of human plasma [17] [14].
- Iron limitation and overall nutrient availability [17].
These conditions mimic the in vivo environment of an infected device, where S. aureus often switches to a biofilm mode of growth, even if it is biofilm-negative under standard in vitro conditions [10].
Prevalence in Clinical Isolates and Correlation with Antibiotic Resistance
The icaADBC operon is highly prevalent in clinical isolates, particularly in those associated with medical device infections. Epidemiological studies reveal a strong correlation between the presence of this operon, multidrug resistance, and treatment failure.
Table 2: Prevalence of icaADBC Genes and Biofilm Formation in Clinical S. aureus Isolates
| Characteristic | Prevalence in Clinical S. aureus Isolates | Significance/Correlation |
|---|---|---|
| icaA | 90% (90/100 isolates) [15] | A core gene for PIA synthesis. |
| icaB | 92% (92/100 isolates) [15] | Essential for PIA maturation and adhesion. |
| icaC | 92% (92/100 isolates) [15] | Critical for PIA export. |
| icaD | 94% (94/100 isolates) [15] | Co-essential with icaA for full activity. |
| MDR Isolates | 100% prevalence of ica genes [15] | Strong link between biofilm formation and multidrug resistance. |
| MRSA Isolates | 96.55% (icaA), 89.65% (icaB/C), 96.55% (icaD) [15] | Highlights biofilm's role in persistent healthcare-associated infections. |
| Therapeutic Failure | 95.4% in IS256+ S. epidermidis infections [13] | IS256-mediated phase variation complicates treatment. |
A study on pediatric device-related infections further underscores the clinical importance of the operon, showing that the presence of icaA and icaD genes along with IS256 was associated with therapeutic failure in most coagulase-negative staphylococcal infections [13]. These findings solidify the icaADBC operon as a key biomarker for virulent, persistent biofilm-forming strains.
Experimental Methodologies for Analysis
Rigorous and standardized experimental protocols are essential for investigating the icaADBC operon and its phenotypic outputs. The following are key methodologies used in the field.
Genotypic Detection by PCR
Purpose: To detect the presence of the icaA, icaD, icaB, and icaC genes, and the insertion sequence IS256. Protocol Summary [15] [13]:
- DNA Extraction: Purify chromosomal DNA from bacterial isolates using a commercial kit (e.g., Promega Wizard Genomic DNA Purification Kit).
- PCR Amplification: Set up 25-50 µL reactions containing PCR buffer, dNTPs, specific primers for each gene, DNA template, and Taq polymerase.
- Cycling Conditions:
- Denaturation: 95°C for 25 seconds.
- Annealing: 52-60°C (gene-specific) for 30 seconds.
- Extension: 72°C for 30 seconds.
- Repeat for 30 cycles, with a final extension at 72°C for 5 minutes.
- Analysis: Analyze PCR products by agarose gel electrophoresis (1.5-2%) and visualize under UV light. Use reference strains (e.g., S. epidermidis RP62A as positive control) for comparison.
Phenotypic Biofilm Assay (Microtiter Plate)
Purpose: To quantitatively assess the biofilm-forming capacity of bacterial isolates [10] [15]. Protocol Summary:
- Inoculation: Dilute an overnight bacterial culture to an OD~600~ of 0.05 in Tryptic Soy Broth (TSB) supplemented with 1% glucose. Add 200 µL per well of a 96-well flat-bottom polystyrene microtiter plate. Include triplicates for each strain and uninoculated broth as a negative control.
- Incubation: Incubate the plate statically for 20-24 hours at 37°C.
- Washing and Staining: Gently remove the planktonic cells and wash the wells twice with 0.15 M phosphate-buffered saline (PBS). Air-dry the plate and stain the adherent biofilm with 0.1% crystal violet for 30 minutes.
- Destaining and Quantification: Wash off excess stain and destain the bound crystal violet with 33% glacial acetic acid (200 µL/well). Measure the absorbance of the eluted dye at 492 nm or 595 nm using a microplate reader. An OD value ≥ 0.17 is typically considered indicative of a biofilm producer.
PIA Detection by Immunofluorescence
Purpose: To visually confirm the production and localization of PIA on bacterial cells in vitro or in ex vivo samples [10]. Protocol Summary:
- Sample Preparation: Grow bacteria on chamber slides anaerobically in TSB with 1% glucose for 48 hours to induce PIA. For in vivo samples, fix exudates from infected tissue cages with paraformaldehyde and spot onto poly-L-lysine-coated slides.
- Fixation and Permeabilization: Fix the samples with 4% formaldehyde for 10 minutes at room temperature.
- Staining: Wash slides with PBS-Tween and incubate with a primary antibody specific for PIA/PNAG. After washing, incubate with a fluorescently labeled secondary antibody.
- Visualization: Examine the slides under a fluorescence microscope. PIA-positive strains will exhibit fluorescent staining surrounding the bacterial cells.
Diagram 2: Experimental Workflow. A flowchart outlining the key methodological steps for genotyping and phenotyping icaADBC-mediated biofilm formation.
The Scientist's Toolkit: Key Research Reagents
The table below catalogs essential reagents and materials required for experimental research on the icaADBC operon and PIA, as derived from established methodologies.
Table 3: Essential Research Reagents for icaADBC and Biofilm Research
| Reagent/Material | Specific Example | Experimental Function |
|---|---|---|
| PCR Primers | icaA-F: GAGGTAAAGCCAACGCACTC [15] | Specific amplification of target genes for genotyping. |
| Growth Media | Tryptic Soy Broth (TSB) with 1% glucose [15] [18] | Supports bacterial growth and induces ica operon expression. |
| Biofilm Assay Plates | 96-well flat-bottom polystyrene microtiter plates [15] | Provides a standardized surface for static biofilm formation. |
| Staining Reagents | 0.1% Crystal Violet solution [15] | Stains adherent bacterial cells in the biofilm for quantification. |
| PIA Extraction Kits | Commercial DNA/Polysaccharide extraction kits | Purifies PIA for structural analysis (e.g., FTIR, NMR) [18]. |
| Anti-PIA/PNAG Antibodies | Specific polyclonal or monoclonal antibodies [18] | Detects PIA production via immunofluorescence or ELISA. |
| Reference Strains | S. epidermidis 1457 (ica+), S. epidermidis M-10/delta ica (ica-) [18] | Essential positive and negative controls for all experiments. |
Beyond Polysaccharides: PIA-Independent Biofilm Mechanisms
While the icaADBC operon is a central pathway, it is not the sole mechanism for biofilm formation in staphylococci. Many clinical isolates form robust biofilms through protein-dependent mechanisms, a strategy often termed PIA-independent biofilm formation [14] [11]. This is mediated by a repertoire of surface proteins known as Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSCRAMMs). Key players include:
- Fibrinogen-binding protein (Fbe/SdrG) and the extracellular matrix-binding protein (Embp), which are highly prevalent in biofilm-positive clinical isolates [14].
- Biofilm-associated protein (Bap), which promotes adhesion and intercellular aggregation via an amyloid fiber-based mechanism [11].
This functional redundancy underscores the adaptability of staphylococci and highlights the need for therapeutic strategies that target both polysaccharide and protein components of the biofilm matrix.
The icaADBC operon remains the definitive genetic blueprint for PIA biosynthesis, playing a non-redundant role in the pathogenesis of staphylococcal device-related infections. Its complex regulation by environmental signals and phase variation mechanisms like IS256 insertion makes it a dynamic and challenging virulence factor. Future research should focus on:
- Developing Anti-Biofilm Therapeutics: Exploiting the knowledge of the ica operon to design inhibitors targeting IcaA enzyme activity or PIA deacetylation by IcaB.
- Immunotherapeutic Approaches: Utilizing PIA as a target for opsonic antibodies, which have shown promise in enhancing bacterial clearance and inhibiting biofilm formation in models of E. coli and staphylococcal infection [18].
- Dual-Targeting Strategies: Creating combination therapies that disrupt both PIA-dependent and MSCRAMM-dependent biofilm pathways to overcome the redundancy in staphylococcal adhesion mechanisms.
A deep understanding of the icaADBC operon is therefore not only fundamental to bacterial genetics but also paves the way for innovative clinical interventions against chronic biofilm-associated infections.
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), serves as a critical exopolysaccharide within the extracellular matrix of staphylococcal biofilms, contributing significantly to the chronicity and irreducibility of device-related infections [3] [1]. This cationic, partially deacetylated polymer of β-1-6-linked N-acetylglucosamine is synthesized by a wide range of bacterial pathogens but represents the dominant exopolysaccharide in staphylococci [3]. Its production is strongly associated with virulence, enabling mechanisms such as immune evasion, resistance to antimicrobial peptides, and protection from phagocytosis [3]. The biosynthesis of PIA is mediated by the icaADBC operon, with each gene encoding enzymes that perform distinct, coordinated functions in the synthesis, modification, and export of this key virulence determinant [3] [1] [2]. Understanding the precise enzymatic functions of IcaA, IcaD, IcaB, and IcaC provides foundational knowledge for developing anti-biofilm strategies targeting serious infections caused by Staphylococcus aureus and Staphylococcus epidermidis [3] [19].
The IcaADBC Operon: Genetic Foundation for PIA Biosynthesis
The ica (intercellular adhesion) locus consists of the biosynthetic operon icaADBC and its regulatory gene, icaR [3] [1]. This locus was first described in S. epidermidis and later found in S. aureus and other staphylococcal species with significant sequence conservation [3] [20]. The icaADBC operon is present in nearly all S. aureus strains and is widespread in clinically significant S. epidermidis isolates, particularly those from device-related infections [3] [19]. Expression of this operon is affected by various environmental conditions and regulated by multiple genetic regulatory systems, contributing to the adaptability of staphylococci in different host environments [3] [1]. Within this operon, each gene encodes a protein with a specific function that collectively enables the production of functional PIA, as detailed in Table 1.
Table 1: Core Enzymatic Components of the icaADBC Operon in PIA Biosynthesis
| Gene | Protein Function | Key Functional Partners | Catalytic Activity/Process |
|---|---|---|---|
| IcaA | N-acetylglucosaminyltransferase [1] [2] | IcaD [1] [2] | Synthesizes PIA oligomers from UDP-N-acetylglucosamine substrate [1] |
| IcaD | Essential cofactor [1] [2] | IcaA [1] [2] | Optimizes efficiency of IcaA transferase activity [1] |
| IcaB | Extracellular de-N-acetylase [1] [5] | Mature PIA polymer [5] | Partially deacetylates PIA, essential for surface retention and biofilm formation [1] [5] |
| IcaC | Putative O-succinyltransferase/Polymer exporter [1] [21] | Nascent polysaccharide chain [1] [21] | Externalizes nascent polysaccharide; potential O-succinylation role [1] [21] |
Enzymatic Functions and Molecular Mechanisms
IcaA: The Central N-acetylglucosaminyltransferase
IcaA functions as the primary N-acetylglucosaminyltransferase responsible for synthesizing the PIA backbone [1] [2]. This enzyme utilizes UDP-N-acetylglucosamine as its substrate to catalyze the formation of β-1,6-glycosidic linkages, thereby creating linear oligomers of N-acetylglucosamine [1]. IcaA is predicted to contain multiple transmembrane domains with a large cytosolic family 2 glycosyltransferase domain, positioning it to utilize cytoplasmic sugar nucleotide precursors while translocating the growing polymer across the membrane [5]. The activity of IcaA alone produces only short oligomers; however, its efficiency is dramatically enhanced through interaction with its essential partner, IcaD [1] [2].
IcaD: The Essential Cofactor for Optimal Polymerization
IcaD is a small integral membrane protein that significantly increases PIA biosynthesis when co-expressed with IcaA [1] [5]. While IcaD possesses no catalytic activity itself, it forms a functional complex with IcaA, granting the optimal efficiency necessary for producing the full-length PIA polymer [1] [2]. This partnership between IcaA and IcaD represents a crucial regulatory checkpoint in PIA synthesis, ensuring that substantial polymer production occurs only when both proteins are expressed, thereby potentially conserving cellular resources until biofilm formation is environmentally advantageous [1].
IcaC: The Putative Transporter and Modifier
IcaC is an integral membrane protein initially proposed to facilitate the externalization of the nascent polysaccharide [1] [21]. Recent bioinformatic analyses have revealed that IcaC contains 10 transmembrane helices and belongs to a large acyltransferase family, suggesting an additional role in the O-succinylation of PIA during biosynthesis [21]. This revised understanding positions IcaC as a bifunctional protein involved in both the transport and chemical modification of the exopolysaccharide. The limited distribution of icaC to the Staphylococcaceae family and its membership in this acyltransferase family indicate that IcaC may represent a staphylococcus-specific therapeutic target [21].
IcaB: The Key Virulence-Activating Deacetylase
IcaB is an extracellular de-N-acetylase that performs the crucial final step in producing functional PIA [1] [5]. As a member of the family 4 carbohydrate esterases (CE4s), IcaB partially deacetylates the PIA polymer by removing acetyl groups from N-acetylglucosamine residues [5]. This partial deacetylation (approximately 15-20% of residues) converts the neutral PIA into a positively charged polymer, which is essential for its retention on the negatively charged bacterial cell surface and for subsequent biofilm accumulation [3] [1]. Strains with deleted icaB genes cannot deacetylate PIA, fail to form biofilms in vitro, and exhibit highly attenuated virulence in animal infection models, underscoring this enzyme's critical role in pathogenesis [5]. The structure of IcaB reveals a (β/α)7 barrel common to CE4 enzymes and includes a conserved surface-exposed hydrophobic loop that likely anchors the protein to the membrane during polysaccharide biosynthesis [5].
Experimental Analysis of PIA Biosynthesis
Methodologies for Investigating Ica Protein Functions
Research into the enzymatic functions of the Ica proteins employs multidisciplinary approaches spanning molecular genetics, biochemistry, and structural biology. Key experimental protocols include:
In Vitro N-acetylglucosaminyltransferase Assay: This biochemical assay measures the ability of IcaA and IcaD to synthesize PIA oligomers from UDP-N-acetylglucosamine. Membrane fractions containing IcaA and IcaD are incubated with UDP-N-acetylglucosamine, and product formation is detected using anti-PIA antibodies or chromatographic methods [20] [5].
IcaB Deacetylase Activity Measurement: The de-N-acetylase activity of IcaB is quantified using colorimetric assays that detect free amino groups released upon deacetylation of purified PIA or synthetic substrates. Metal dependence is assessed by testing activity in the presence of different divalent cations (Ni²⁺, Co²⁺, Zn²⁺), with maximum deacetylation rates typically observed with these cofactors [5].
Biofilm Formation Assays: The functional outcome of Ica enzyme activities is frequently assessed through biofilm formation assays using methods such as safranin staining in microtiter plates [20] or confocal microscopy analysis of biofilm architecture following genetic manipulation of ica genes [19].
Gene Expression Analysis: Reverse transcription PCR (RT-PCR) and quantitative real-time PCR are used to measure transcription levels of icaADBC genes under different environmental conditions, such as after cold stress [22].
Quantitative Experimental Data on Ica Gene Expression and Function
Table 2: Experimental Data on Ica Gene Expression and Enzymatic Function
| Experimental Approach | Key Findings | Significance/Implication |
|---|---|---|
| Gene Deletion Studies [20] [5] | ΔicaADBC strains: Complete loss of biofilm formation and PIA production [20]. ΔicaB strains: Produce PIA but fail to form biofilms; PIA is released into medium [5]. | Confirms essential role of entire operon; demonstrates IcaB critical for surface retention. |
| In Vitro Enzyme Assays [5] | IcaA + IcaD: Generate PIA-recognized antibodies in vitro. IcaB: Maximum deacetylation with Ni²⁺, Co²⁺, Zn²⁺. | Demonstrates sufficiency of IcaA/D for polymerization; reveals IcaB metal dependence. |
| Transcriptional Analysis [22] | Cold stress (20 weeks, -20°C): >2-fold increase in icaA, icaB, icaC, icaD mRNA in multiple S. aureus strains. | Environmental stress modulates ica operon expression; may enhance biofilm potential. |
| Structural Studies [5] | IcaB structure: (β/α)7 barrel with circularly permuted CE4 motifs; conserved hydrophobic loop for membrane association. | Provides basis for catalytic mechanism understanding; suggests inhibitor design strategies. |
Visualization of the PIA Biosynthesis Pathway
The following diagram illustrates the coordinated biosynthetic pathway for PIA synthesis and export, highlighting the specialized roles of each Ica protein:
Diagram 1: PIA Biosynthesis and Export Pathway. This schematic illustrates the sequential enzymatic process where IcaA and IcaD synthesize the PNAG polymer from UDP-GlcNAc precursors, IcaC facilitates export and potential modification, and IcaB's deacetylation creates the positively charged PIA essential for biofilm formation.
Research Reagents and Methodological Toolkit
Table 3: Essential Research Reagents for Investigating Ica Enzymes and PIA Biosynthesis
| Reagent/Cell Line | Specific Application | Experimental Function/Rationale |
|---|---|---|
| S. epidermidis RP62A (ATCC 35984) [20] | Reference strain for PIA studies | Strong biofilm producer; contains functional icaADBC operon; used as positive control. |
| S. aureus SA113 (ATCC 35556) [20] | S. aureus model for PIA research | Derived from NCTC 8325; restriction deficient; well-characterized ica operon. |
| Anti-PIA Antibody [20] | PIA detection and quantification | Specifically recognizes PIA/PNAG epitopes; used in Western blot, ELISA, and immunofluorescence. |
| Proteinase K [22] [20] | Biofilm matrix disruption | Degrades protein components in biofilm; distinguishes PIA-dependent vs protein-dependent biofilms. |
| Dispersin B [2] | PIA-specific degradation | Glycoside hydrolase that cleaves β-1,6-GlcNAc linkages; confirms PIA presence in biofilm matrix. |
| Congo Red Agar (CRA) [2] | Phenotypic biofilm screening | Differentiates biofilm-producing (black colonies) from non-producing strains (red colonies). |
| icaADBC Deletion Mutants [20] [5] | Functional genetic studies | Isogenic mutants (ΔicaA, ΔicaB, etc.) essential for establishing gene-specific functions. |
Research Implications and Therapeutic Applications
Understanding the precise enzymatic functions of the Ica proteins has direct implications for developing novel anti-biofilm strategies. The essential role of PIA in staphylococcal biofilm formation and immune evasion, combined with its conservation across many pathogenic species, makes it an attractive target for vaccines and inhibitory compounds [3] [1]. The partially deacetylated form of PNAG has shown promise as a vaccine candidate, inducing protective antibodies that promote opsonophagocytosis [1] [2]. Enzyme-based approaches targeting PIA include Dispersin B, which degrades the polysaccharide backbone, and investigational compounds that inhibit IcaB's deacetylase activity [2]. The staphylococcus-specific distribution of IcaC further identifies this protein as a potential target for narrow-spectrum anti-biofilm agents that would minimally impact commensal flora [21].
In conclusion, the coordinated activities of IcaA, IcaD, IcaB, and IcaC enable the production, modification, and export of PIA, a fundamental determinant of staphylococcal biofilm formation and pathogenesis. Continued research into the structural and mechanistic details of these enzymes will undoubtedly yield new therapeutic approaches to combat device-related and other chronic bacterial infections.
Environmental Regulation and Phase Variation of PIA Expression
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), serves as a fundamental component of the extracellular matrix in staphylococcal biofilms. This cationic, partially deacetylated polymer of β-1-6-linked N-acetylglucosamine confers critical functional properties including intercellular adhesion, protection from host immune defenses, and enhanced antibiotic resistance [3] [2]. The biosynthesis of PIA is mediated by the icaADBC operon, with IcaA and IcaD synthesizing short oligomers, IcaC facilitating polymerization and translocation, and IcaB responsible for the partial deacetylation essential for its cationic nature and adherence properties [3] [2]. Understanding the sophisticated regulatory mechanisms controlling PIA expression—particularly environmental regulation and phase variation—is paramount for developing effective anti-biofilm strategies against Staphylococcus aureus and Staphylococcus epidermidis, the leading etiologic agents of implant-associated infections [23] [2].
Molecular Mechanisms of Phase Variation in PIA Expression
Phase variation represents a reversible, high-frequency switch in gene expression that provides bacterial populations with a bet-hedging strategy to adapt to fluctuating environments. In staphylococci, this phenomenon manifests as reversible ON/OFF switching of PIA production, mediated by distinct genetic mechanisms in S. epidermidis and S. aureus.
Insertion Sequence Element IS256-Mediated Phase Variation inS. epidermidis
In S. epidermidis, phase variation occurs primarily through the reversible insertion and excision of the insertion sequence element IS256 within the ica operon. Molecular analyses of biofilm-negative variants revealed that approximately 30% of these variants lost PIA production due to IS256 transposition into either the icaA or icaC genes, effectively disrupting operon functionality [24]. Crucially, this transposition event is reversible; upon repeated passages of PIA-negative insertional mutants, the biofilm-forming phenotype was restored through complete excision of IS256, including the initially duplicated 8 bp target sites [24]. This reversible insertion mechanism represents a dedicated molecular switch for virulence modulation in S. epidermidis.
Slipped-Strand Mispairing inS. aureus
In contrast, S. aureus employs a different mechanism involving slipped-strand mispairing within a simple tetranucleotide tandem repeat (TTTA) naturally located within the icaC gene [25]. During DNA replication, mispairing between mother and daughter strands in this repetitive region results in the addition or subtraction of repeat units. When the number of inserted or deleted repeats is not a multiple of three, the translational reading frame shifts, producing a truncated, non-functional IcaC protein [25]. This reversible, RecA-independent process serves as a phase variation mechanism, with PIA-negative variants demonstrating a fitness advantage under certain conditions, suggesting that PIA production carries a metabolic cost that may be disadvantageous in specific environments [25].
Table 1: Comparative Mechanisms of Phase Variation in Staphylococci
| Characteristic | S. epidermidis | S. aureus |
|---|---|---|
| Molecular Mechanism | Reversible insertion/excision of IS256 element | Slipped-strand mispairing in tandem repeats |
| Genetic Target | icaA or icaC genes | Tetranucleotide (TTTA) repeats within icaC |
| Frequency | Observed in ~30% of biofilm-negative variants | Most frequent mutation in non-mucoid variants |
| Reversibility | Complete excision of IS256 with target sites | Expansion/contraction of repeats |
| Regulatory Dependence | Independent of RecA | RecA-independent |
| Biological Consequence | Frameshift or disruption of ica genes | Frameshift mutation truncating IcaC |
Environmental Regulation of PIA Expression
PIA biosynthesis responds to diverse environmental signals through complex regulatory networks, enabling staphylococci to optimize biofilm formation for specific ecological niches.
Key Environmental Regulators
Anaerobic Conditions & Oxidative Stress: Anaerobiosis serves as a potent inducer of ica operon expression and PIA production in both S. epidermidis and S. aureus [23]. This response is partially mediated through the alternative sigma factor σB (SigB), a global regulator of the general stress response [26] [2]. The σB-dependent activation demonstrates strain-specific characteristics, as evidenced by contrasting biofilm responses to diosgenin treatment in different S. aureus strains [26].
Osmolarity and Substrate Availability: High salt concentrations (4% NaCl) and the presence of glucose significantly enhance biofilm formation by activating ica expression [18] [23]. Additionally, oleic acid induces S. aureus biofilm formation, likely through ionic interactions between the positively charged PIA and negatively charged fatty acids [23].
Temperature and pH Fluctuations: Environmental temperature shifts and pH variations influence the stability of tandem repeats in icaC and affect the activity of regulatory systems controlling the ica operon, though these factors require further characterization [2].
Quorum-Sensing Systems
The accessory gene regulator (agr) quorum-sensing system typically functions as a negative regulator of biofilm formation by repressing the expression of surface-associated adhesins, including PIA [2]. As bacterial density increases and Agr activity peaks, the system promotes a dispersal phase that facilitates dissemination to new infection sites [2]. This counterintuitive regulation creates a dynamic equilibrium between biofilm formation and dispersal throughout the infection cycle.
Global Regulatory Network
The staphylococcal accessory regulator A (SarA) activates icaADBC transcription by binding to the operon's promoter region [23]. SarA expression itself is activated by σB, establishing a key regulatory circuit: σB → SarA → icaADBC [26]. Additionally, the SaeRS two-component system responds to environmental stimuli such as neutrophil-induced stressors and modulates ica expression, though its impact varies substantially between strains [17].
Experimental Approaches for Studying PIA Regulation
Assessing Phase Variation and Biofilm Phenotypes
Congo Red Agar (CRA) Plate Assay: This phenotypic screening method distinguishes PIA-producing and non-producing variants based on colony morphology. PIA-producing colonies typically appear dry with irregular edges, while PIA-negative variants form slick, circular colonies, often surrounded by a transparent red perimeter [25].
In Vitro Biofilm Formation Assay: A standard microtiter plate protocol involves several key steps [18]:
- Adjust bacterial culture optical density (OD₆₀₀) to 0.7
- Dilute cultures 1:200 in Brain Heart Infusion Broth supplemented with 1% glucose (BHIGlc) and 4% NaCl (BHINaCl)
- Transfer 200 μL aliquots to polystyrene microtiter plates
- Incubate static for 24 hours at 37°C
- Remove planktonic cells and stain adherent biomass with 1% crystal violet
- Dissolve bound stain in 30% acetic acid and measure absorbance at 595nm
Molecular Analysis of Phase Variants: For genetic characterization, Southern hybridization with ica-specific probes, polymerase chain reaction (PCR) amplification of the ica locus, and nucleotide sequencing identify IS256 insertion sites or tandem repeat variations [24] [25].
Monitoring PIA Biosynthesis and Regulation
PIA Extraction and Purification: A standardized protocol for PIA isolation includes [27] [18]:
- Culture bacteria in Tryptic Soy Broth with gentle agitation (40-50 rpm) for 24 hours
- Harvest cells by centrifugation at 1000 × g for 20 minutes
- Resuspend in PBS (pH 7.5) and disrupt by sonication
- Purify PIA using size exclusion chromatography (Fast Protein Liquid Chromatography)
- Confirm PIA identity via colorimetric assays, Fourier-transform infrared spectroscopy, and proton nuclear magnetic resonance spectroscopy
Gene Expression Analysis: Quantitative real-time PCR monitors changes in icaADBC transcription under different environmental conditions. For example, diosgenin treatment revealed strain-specific expression patterns, with decreased icaA and spa expression correlating with reduced biofilm formation in some strains [26].
Table 2: Key Methodologies for Investigating PIA Regulation
| Methodology | Application | Key Outcomes |
|---|---|---|
| Congo Red Agar Screening | Rapid phenotyping of phase variants | Distinguishes PIA-positive (dry, irregular) from PIA-negative (slick, circular) colonies |
| Microtiter Biofilm Assay | Quantitative biofilm assessment | Spectrophotometric measurement (OD₅₉₅) of adhered biomass |
| Size Exclusion Chromatography | PIA purification and analysis | Determines molecular weight distribution near void volume |
| Southern Hybridization/PCR | Genetic analysis of phase variants | Identifies IS256 insertions or tandem repeat variations in ica locus |
| qRT-PCR | Gene expression profiling | Quantifies changes in icaADBC transcription under different conditions |
| FTIR and NMR Spectroscopy | Structural confirmation of PIA | Verifies chemical identity as poly-β(1-6)-N-acetylglucosamine |
Research Reagent Solutions Toolkit
Table 3: Essential Research Reagents for PIA and Biofilm Studies
| Reagent / Material | Function / Application | Experimental Context |
|---|---|---|
| Polystyrene Microtiter Plates | Substrate for in vitro biofilm formation | Standardized assessment of biofilm formation capacity [18] |
| Tryptic Soy Broth (TSB) | Growth medium for PIA production | Supports optimal PIA yield under gentle agitation [18] |
| Brain Heart Infusion Broth with 1% Glucose & 4% NaCl (BHIGlc/BHINaCl) | Biofilm-inducing medium | Enhances ica expression and biofilm formation in assays [18] |
| Congo Red Agar (CRA) | Differential medium for PIA phenotyping | Visual identification of PIA-producing and phase variants [25] |
| Anti-PIA/PNAG Antibodies | Detection, quantification, and functional studies | Measures opsonophagocytosis (40.48% lethality in E. coli) and biofilm inhibition [18] |
| Diosgenin (Plant Steroid) | Chemical probe for regulatory studies | Elucidates σB-dependent regulatory pathways through strain-specific effects [26] |
| Fast Protein Liquid Chromatography (FPLC) | PIA purification | Size-exclusion chromatography for isolating PIA from bacterial extracts [27] |
Regulatory Pathways and Experimental Framework
The following diagram illustrates the complex regulatory network controlling PIA expression and the experimental approaches for its investigation:
Diagram 1: Integrated regulatory network of PIA expression. Environmental signals converge on regulatory systems that control the icaADBC operon. Phase variation occurs through IS256 insertion (S. epidermidis) or tandem repeat changes (S. aureus). Experimental methods detect each component of the system.
The environmental regulation and phase variation of PIA expression represent sophisticated adaptive mechanisms that enhance staphylococcal persistence in hostile environments, including during human infection. The dual-layered control system—comprising rapid, reversible phase variation and finely-tuned environmental responsiveness—enables bacterial populations to maintain phenotypic heterogeneity, optimizing survival across fluctuating conditions. Understanding these mechanisms at molecular depth provides crucial insights for developing targeted anti-biofilm strategies, such as quorum-quenching approaches that disrupt dispersal signals or immunotherapeutic interventions leveraging PIA-specific antibodies. Future research elucidating the precise environmental sensing mechanisms and the fitness trade-offs governing phase variation will unlock new opportunities for combating biofilm-associated infections, particularly those involving indwelling medical devices where staphylococci predominate.
Polysaccharide intercellular adhesin (PIA), a poly-β(1-6)-N-acetylglucosamine (PNAG) partially deacetylated, is a crucial component of the extracellular polymeric substance of staphylococcal biofilms [28]. This whitepaper examines the phylogenetic distribution of PIA and its homologs across bacterial species, with particular focus on implications for biofilm-related pathogenesis and therapeutic interventions. The presence of PNAG exopolysaccharides in diverse bacteria indicates an evolutionarily recurrent strategy for biofilm formation and immune evasion. Understanding this distribution provides critical insights for developing broad-spectrum anti-biofilm strategies and vaccines targeting this conserved virulence factor.
PIA/PNAG Structure and Function in Staphylococci
In Staphylococcus aureus and Staphylococcus epidermidis, PIA is a key virulence factor responsible for biofilm accumulation on implanted medical devices [28]. The synthesis of PIA is mediated by the icaADBC locus, where:
- IcaA: N-acetylglucosaminyltransferase that synthesizes PIA oligomers
- IcaD: Enhances the efficiency of IcaA
- IcaC: Involved in externalization of the polysaccharide
- IcaB: N-deacetylase responsible for partial deacetylation of PIA [28]
The deacetylation of PIA generates a positively charged polymer that facilitates intercellular adhesion and confers resistance to host immune defenses and antibiotic treatments [28]. Bacteria in biofilms can resist antibiotics at concentrations up to 1000 times higher than those effective against planktonic cells [28].
Phylogenetic Distribution of PIA/PNAG Homologs
PNAG in Gram-Negative Bacteria
Recent research has confirmed that Escherichia coli produces an exopolysaccharide biochemically indistinguishable from staphylococcal PIA, known as poly-β(1,6)-N-acetylglucosamine (PNAG) [18]. This PNAG exopolysaccharide in E. coli is synthesized through the pgaABCD operon and enhances bacterial adhesion capabilities, promotes cell-to-cell aggregation, stabilizes the biofilm matrix, and provides protection against environmental stresses and antimicrobial agents [18].
Table 1: Bacterial Species Producing PIA/PNAG Exopolysaccharides
| Bacterial Species | Gram Reaction | Exopolysaccharide | Genetic Locus | Key Functions |
|---|---|---|---|---|
| Staphylococcus epidermidis | Positive | PIA | icaADBC | Biofilm formation, immune evasion |
| Staphylococcus aureus | Positive | PIA | icaADBC | Biofilm formation, immune evasion |
| Escherichia coli | Negative | PNAG | pgaABCD | Surface adhesion, biofilm matrix stability |
| Streptococcus pneumoniae | Positive | (Capsular) | pia | Iron acquisition (hydroxamate siderophore transport) |
It is noteworthy that Streptococcus pneumoniae possesses a pia gene, though this encodes a substrate-binding protein for iron acquisition rather than a polysaccharide adhesin [29]. This highlights the importance of distinguishing between genetically similarly named but functionally distinct elements.
Conservation and Evolutionary Implications
The structural and functional conservation of PIA/PNAG across Gram-positive and Gram-negative bacteria suggests this exopolysaccharide represents an evolutionarily optimized strategy for biofilm formation and persistence in hostile environments. The presence of biochemically similar polysaccharides in diverse pathogens indicates convergent evolution or horizontal gene transfer of this virulence trait.
Experimental Evidence for Cross-Species PNAG Functionality
Antibody Cross-Reactivity and Biofilm Inhibition
A recent investigation demonstrated that antibodies generated against staphylococcal PIA can bind to E. coli PNAG and significantly impact biofilm formation [18]. Key experimental findings include:
- Purified PIA from S. epidermidis induced protective antibodies with 40.48% opsonization properties against E. coli [18]
- Sera from PIA-immunized groups showed significant increases in antibody production and protective IgG titer levels compared to control groups [18]
- Anti-PIA antibodies demonstrated substantial biofilm inhibition in E. coli in vitro compared to non-immunized serum [18]
These findings confirm the structural and antigenic similarity between staphylococcal PIA and E. coli PNAG, suggesting potential for cross-species therapeutic targeting.
PIA Extraction and Purification Protocol
The following detailed methodology has been established for PIA extraction and purification [18]:
- Bacterial Culture: Grow bacterial strains in 2 liters of Tryptic Soy Broth (TSB) at 37°C with gentle agitation (40-50 rpm/min) for 24 hours
- Harvesting: Collect cells by centrifugation at 1000 × g for 20 minutes at 4°C
- Resuspension: Resuspend cell pellet in 20 mL of PBS (pH 7.5)
- Sonication: Sonicate four times for 30 seconds each on ice
- Centrifugation: Centrifuge at 12,000 rpm for 15 minutes
- Purification: Further purification steps as required for experimental application
The extracted polysaccharide can be confirmed using FTIR, NMR, and colorimetric methods [18].
In Vitro Biofilm Formation Assay
Standardized protocol for assessing biofilm formation [18]:
- Adjust the optical density (OD₆₀₀) of inoculated colonies in BHI broth to 0.7
- Dilute supernatants 1:200 in BHI broth enriched with 1% (w/w) D-glucose (BHIGlc) and 4% (w/w) NaCl (BHINaCl)
- Transfer 200 μL of diluted supernatants to polystyrene microtiter plates
- Incubate for 24 hours at 37°C
- Remove planktonic cells by washing each well three times with phosphate-buffered saline (PBS)
- Add 150 μL of 1% crystal violet solution to each well
- Dissolve stain bound to adherent cells in 160 μL of 30% acetic acid
- Measure absorbance spectrophotometrically at 595 nm
Visualization of PIA/PNAG Phylogenetic Distribution and Functional Relationships
PIA and PNAG Phylogenetic Distribution
This diagram illustrates the phylogenetic distribution of PIA/PNAG across bacterial species and demonstrates the functional consequences of antibody cross-reactivity, highlighting the potential for broad-spectrum anti-biofilm strategies.
The Scientist's Toolkit: Essential Research Reagents
Table 2: Key Research Reagents for PIA/PNAG Studies
| Reagent/Solution | Function | Application Example |
|---|---|---|
| Tryptic Soy Broth (TSB) | Bacterial culture medium | PIA production in S. epidermidis [18] |
| Phosphate-Buffered Saline (PBS) | Buffer for biological reactions | Cell resuspension during PIA extraction [18] |
| Crystal Violet Solution (1%) | Biofilm staining | Quantitative biofilm formation assay [18] |
| BHIGlc (BHI + 1% Glucose) | Biofilm-promoting medium | Enhanced biofilm formation in vitro [18] |
| BHINaCl (BHI + 4% NaCl) | Osmotic stress medium | Induction of biofilm formation [18] |
| Anti-PIA/PNAG Antibodies | Immunological detection and functional inhibition | Opsonization and biofilm disruption studies [18] |
| Acetic Acid (30%) | Solvent for crystal violet | Dissolution of stained biofilms for quantification [18] |
| Mueller Hinton Agar | Antibiotic susceptibility testing | Confirmation of transmutant strains [18] |
Therapeutic Implications and Future Directions
The phylogenetic distribution of PIA/PNAG has significant implications for therapeutic development. The cross-reactive nature of anti-PIA antibodies suggests potential for broad-spectrum approaches targeting biofilms across multiple bacterial species [18]. Two primary strategic approaches are emerging:
- Biofilm Disaggregation: Utilizing enzymes such as Dispersin B (targets PNAG) or DNase I (targets extracellular DNA) to disrupt established biofilms [28]
- Quorum Quenching: Interfering with bacterial communication systems to prevent biofilm formation [28]
The partial deacetylation of PNAG creates a potential epitope for vaccine development, as deacetylated PNAG retained on the surface of S. aureus favors opsonophagocytosis and represents a candidate for immune protection [28]. Further research is needed to explore the extent of PIA/PNAG homologs across additional bacterial species and optimize cross-reactive therapeutic agents.
Investigating PIA: Analytical Techniques and Therapeutic Applications
PIA Extraction and Purification Protocols from Staphylococcal Cultures
Polysaccharide Intercellular Adhesin (PIA) is a crucial exopolysaccharide component of the extracellular matrix in staphylococcal biofilms, particularly in Staphylococcus epidermidis and Staphylococcus aureus [1] [3]. Chemically, PIA is a linear polymer of β-1,6-linked N-acetylglucosamine (poly-β(1,6)-N-acetylglucosamine or PNAG) that is partially deacetylated (approximately 15-20%), giving it a positive charge [3] [2]. This cationic nature contributes significantly to its role in bacterial adhesion, biofilm matrix stability, and immune evasion mechanisms [3].
The biomedical significance of PIA extraction extends beyond basic research. As a conserved pan-pathogen surface polysaccharide, PIA is a promising target for therapeutic and vaccine development [6]. The presence of PIA is strongly associated with virulence in device-related infections, and antibodies against PIA have demonstrated protective effects in animal models [18] [30] [31]. Purified PIA serves as a critical antigen for generating specific antibodies that can inhibit biofilm formation and promote opsonophagocytic killing of biofilm-forming bacteria [30] [6].
PIA Biosynthesis and Genetic Regulation
PIA biosynthesis in staphylococci is mediated by the icaADBC operon (intercellular adhesion) [1] [3]. Each gene in this operon plays a distinct role: IcaA is an N-acetylglucosaminyltransferase that synthesizes PIA oligomers from UDP-N-acetylglucosamine; IcaD enhances the efficiency of IcaA; IcaC is involved in externalizing the nascent polysaccharide; and IcaB is an N-deacetylase responsible for the partial deacetylation that is critical for PIA function [1] [3]. The expression of this operon is influenced by environmental conditions and regulated by various genetic and metabolic factors [1] [2].
The following diagram illustrates the biosynthetic pathway and extraction workflow for PIA:
Detailed Extraction and Purification Methodology
Bacterial Cultivation and Initial Processing
For optimal PIA production, standard strains of S. epidermidis (such as 1457) should be cultivated in Tryptic Soy Broth (TSB) under conditions that promote biofilm formation [18] [30] [31]. The specific protocol is as follows:
- Inoculation and Incubation: Grow bacteria in 2 liters of TSB at 37°C for 24 hours with gentle agitation (40-50 rpm/min) to promote PIA production without excessive shear stress [18] [30] [31].
- Cell Harvesting: Centrifuge the culture at 1000 × g for 20 minutes at 4°C to pellet bacterial cells [18] [30].
- Cell Resuspension: Resuspend the harvested cells in 20 mL of phosphate-buffered saline (PBS, pH 7.5) to prepare for macromolecule extraction [18] [30] [31].
Cell Disruption and Primary Extraction
Effective disruption of bacterial cells is essential for releasing PIA into solution while maintaining its structural integrity:
- Sonication: Subject the cell suspension to sonication on ice, typically four times for 30 seconds each, with adequate cooling intervals to prevent overheating [18] [30] [31].
- Primary Clarification: Centrifuge the sonicated suspension at 12,000 rpm for 15 minutes at 4°C to remove cell debris [18] [30].
- Supernatant Collection: Carefully collect the supernatant containing the crude PIA extract [18] [30].
Purification Techniques
The purification process involves multiple steps to isolate PIA from other cellular components:
- Enzymatic Treatment: Incubate the supernatant with proteinase K to digest contaminating proteins [18] [30] [31].
- Dialysis and Concentration: Dialyze the sample against PBS using membranes with a 12 kDa molecular weight cutoff to remove small molecules, then concentrate using Centriprep 10 devices (Amicon) [27] [31].
- Size Exclusion Chromatography: Apply the concentrated sample to a Sephacryl S-100 column (1.6 × 100 cm) equilibrated with 50 mM sodium phosphate buffer [27] [31]. Elute at a flow rate of 1 mL/min, collecting 5 mL fractions over 5-minute intervals [31].
- PIA Identification: PIA typically elutes in fractions near the void volume of the column, indicating high molecular weight polymers [27].
Quantitative Analysis of Purified PIA
Compositional Analysis
Table 1: Biochemical Composition of Purified PIA from S. epidermidis
| Component | Quantitative Measurement | Analytical Method | Functional Significance |
|---|---|---|---|
| Hexosamine | 5700 µg/mL (65% content) | MBTH colorimetric assay [27] [31] | Reflects N-acetylglucosamine backbone |
| Hexose | 620 µg/mL | Phenol-sulfuric acid method [27] | Detects contaminating sugars |
| Ketoses | 170 µg/mL | Colorimetric assay [27] | Monitors purification quality |
| Hemagglutination Titer | 1:128 | Hemagglutination assay [27] | Indicates biological activity |
Structural Validation Techniques
Table 2: Structural Analysis Methods for PIA Characterization
| Analytical Method | Key Findings | Technical Parameters |
|---|---|---|
| Fourier Transform Infrared (FTIR) Spectroscopy | Confirms glycosidic linkages and acetylation patterns [27] [31] | KBr pellet method; 256 scans averaged [31] |
| Nuclear Magnetic Resonance (NMR) Spectroscopy | Verifies β-1,6 linkages and deacetylation pattern [18] [27] | Proton NMR analysis [27] |
| Colorimetric Assays | Quantifies hexosamine content and carbohydrate composition [18] [27] | MBTH method for hexosamine; phenol-sulfuric for total carbs [27] [31] |
Essential Research Reagents and Solutions
Table 3: Key Reagents for PIA Extraction and Purification
| Reagent/Equipment | Specification | Function in Protocol |
|---|---|---|
| Tryptic Soy Broth (TSB) | Standard culture medium | Bacterial growth and PIA induction |
| Phosphate Buffered Saline (PBS) | 50 mM, pH 7.5 | Cell resuspension and buffer system |
| Proteinase K | Molecular biology grade | Degradation of contaminating proteins |
| Sephacryl S-100 | Size exclusion matrix | Chromatographic purification of PIA |
| Centriprep 10 | 10 kDa MWCO | Concentration and buffer exchange |
| Dialysis Membrane | 12 kDa MWCO | Removal of small molecules |
| Sonication Equipment | Probe sonicator with cooling | Cell disruption and PIA release |
Applications in Biofilm Research and Therapeutic Development
Purified PIA has significant applications in both basic research and translational medicine:
Immunological Studies
- Vaccine Development: PIA serves as a protective antigen that induces opsonic and protective antibodies [18] [30]. Studies demonstrate that PIA-immunized mice show significantly increased IgG titers compared to control groups [30] [31].
- Antibody Characterization: Purified PIA enables the development of monoclonal antibodies like F598 (in phase II clinical trials) and TG10, which target different acetylation states of PNAG [6].
- Biofilm Inhibition: Anti-PIA antibodies demonstrate substantial biofilm inhibition in vitro, with studies showing 40.48% opsonic lethality against E. coli expressing PNAG [18] [30].
Structural-Functional Relationships
Recent research utilizing monoclonal antibodies with different PIA binding specificities has revealed that Staphylococcus biofilms contain distinct regions of high and low N-acetylation, rather than polysaccharides with intermittent deacetylation throughout the biofilm [6]. This architectural insight has led to novel therapeutic strategies combining antibodies that target both highly acetylated and deacetylated exopolysaccharides, resulting in 90% survival in a lethal S. aureus challenge murine model [6].
Technical Considerations and Quality Control
Purity Assessment and Contaminant Removal
- Endotoxin Testing: Use Limulus amebocyte lysate assays to quantify endotoxin content, with acceptable levels typically below 4.5 EU/mL for immunological applications [31].
- Enzymatic Cleanup: Treat purified PIA preparations with DNase and RNase to remove contaminating nucleic acids [31].
- Pyrogenicity and Toxicity Testing: Perform in vivo safety assessments through animal inoculation studies [31].
Storage and Stability
- Storage Conditions: Purified PIA should be stored at -20°C in appropriate buffers to maintain stability [18] [27].
- Structural Integrity: Regular validation of structural integrity through FTIR or NMR is recommended for long-term stored samples.
This comprehensive protocol for PIA extraction and purification provides researchers with standardized methods to obtain high-quality PIA for biofilm research, vaccine development, and therapeutic antibody production. The consistent application of these techniques will enhance reproducibility across studies and facilitate advancements in combating biofilm-associated infections.
Polysaccharide Intercellular Adhesin (PIA) is a crucial virulence factor in staphylococcal biofilms, particularly in Staphylococcus epidermidis and Staphylococcus aureus. As the primary substance responsible for bacterial accumulation and biofilm formation on implanted medical devices, PIA enables these pathogens to cause chronic and device-related infections. Structurally, PIA is a linear polymer of β-1,6-linked N-acetylglucosamine (GlcNAc) residues that are partially deacetylated [1] [2]. This partial deacetylation, typically between 10-30% of the residues, creates a positively charged polymer at physiological pH, which enhances its adhesive properties and interaction with bacterial surfaces and host components [1]. The synthesis of PIA is mediated by the icaADBC operon, where the icaA and icaD genes are responsible for polymerization, while icaB encodes the N-deacetylase that creates the partially deacetylated structure [2].
The structural confirmation of PIA presents significant analytical challenges due to its high molecular weight, variable degree of deacetylation, and complex interactions within the biofilm matrix. Researchers must employ a multimodal analytical approach to fully characterize this critical biofilm component. This technical guide provides an in-depth overview of the primary methods—Fourier-Transform Infrared (FTIR) spectroscopy, Nuclear Magnetic Resonance (NMR) spectroscopy, and colorimetric assays—used for the structural confirmation and quantification of PIA within the context of staphylococcal biofilm research. These techniques enable researchers to confirm polymer identity, determine structural modifications, quantify specific functional groups, and establish structure-function relationships critical for understanding biofilm pathogenesis and developing anti-biofilm strategies [32] [30] [33].
Experimental Protocols for PIA Preparation and Analysis
PIA Extraction and Purification Methodology
The structural analysis of PIA requires careful extraction and purification to obtain the native polymer without structural degradation. The following protocol, adapted from established methods in the literature, has been optimized for staphylococcal biofilms [30]:
Bacterial Culture and Biofilm Production: Inoculate Staphylococcus epidermidis (e.g., strain 1457) in 2 liters of Tryptic Soy Broth (TSB) supplemented with 1% (w/w) glucose (BHIGlc) and 4% (w/w) NaCl (BHINaCl) to enhance PIA production. Incubate at 37°C with gentle agitation (40-50 rpm/min) for 24 hours to promote biofilm formation [30].
Harvesting and Initial Processing: Harvest bacterial cells by centrifugation at 1,000 × g for 20 minutes at 4°C. Discard the supernatant and resuspend the cell pellet in 20 mL of phosphate-buffered saline (PBS, pH 7.5). Keep samples on ice throughout the process to prevent enzymatic degradation [30].
Cell Disruption and Crude PIA Isolation: Disrupt the bacterial cells using sonication on ice (four cycles of 30 seconds each with adequate cooling intervals between cycles). Centrifuge the lysate at 12,000 rpm for 15 minutes at 4°C to remove cell debris. Collect the supernatant containing crude PIA and other extracellular materials [30].
Concentration and Preliminary Purification: Concentrate the supernatant using a Centriprep 10 device (Amicon, Witten, Germany) with a 10 kDa molecular weight cut-off to retain PIA while removing smaller contaminants. Alternatively, ultrafiltration devices with similar specifications can be used [30].
Protein Removal and Final Purification: Treat the concentrated sample with proteinase K (0.5-1.0 mg/mL) at 37°C for 2-4 hours to digest contaminating proteins. Subsequently, purify the PIA using a C18 column (KNAUER, Germany) pre-equilibrated with 50 mM PBS to remove residual lipids and other hydrophobic contaminants. The purified PIA can be stored at -20°C or -80°C for long-term preservation [30].
Fourier-Transform Infrared (FTIR) Spectroscopy Analysis
FTIR spectroscopy provides crucial information about the functional groups and chemical bonds present in PIA, serving as a fingerprint for initial structural identification:
Sample Preparation: Prepare purified PIA samples by lyophilization and create pellets using approximately 2 mg of dried PIA mixed with 200 mg of potassium bromide (KBr). Alternatively, for attenuated total reflectance (ATR) FTIR, place a small amount of purified PIA directly onto the ATR crystal [32].
Spectral Acquisition: Acquire FTIR spectra in the range of 4000-400 cm⁻¹ with a resolution of 4 cm⁻¹. Perform 32-64 scans to ensure adequate signal-to-noise ratio. Always collect a background spectrum under identical conditions [32].
Key Spectral Interpretations for PIA: Examine the resulting spectrum for characteristic absorption bands:
- Amide A (~3300 cm⁻¹): N-H stretching vibration
- Amide I (~1650 cm⁻¹): C=O stretching vibration of acetyl groups
- Amide II (~1560 cm⁻¹): N-H bending and C-N stretching vibrations
- C-OH stretching (~1030-1080 cm⁻¹): Carbohydrate backbone vibrations [32]
The degree of N-acetylation can be estimated by comparing the ratio of amide I or amide II bands to carbohydrate reference bands, with calibration against standards of known acetylation degree [32].
Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
NMR spectroscopy, particularly ( ^1H )-NMR and ( ^{13}C )-NMR, provides detailed information about the atomic structure and connectivity of PIA:
Sample Preparation: Dissolve 5-10 mg of purified PIA in 0.5-0.7 mL of deuterated solvent—typically D₂O for native PIA or deuterated dimethyl sulfoxide (DMSO-d₆) for partially acetylated forms. For full structural characterization, exchange labile protons with deuterium by repeatedly dissolving in D₂O and lyophilizing [32] [34].
( ^1H )-NMR Acquisition Parameters:
( ^{13}C )-NMR Acquisition Parameters:
Key NMR Assignments for PIA:
Two-dimensional NMR techniques such as COSY, TOCSY, HSQC, and HMBC can provide additional connectivity information for complete structural elucidation, particularly for confirming the β-(1→6) glycosidic linkage pattern unique to PIA/PNAG [32].
Colorimetric Assays for Functional Group Quantification
Colorimetric assays provide quantitative data on specific functional groups and composition of PIA:
Total Carbohydrate Content - Phenol-Sulfuric Acid Method:
- Reagents: 5% (w/v) aqueous phenol, concentrated sulfuric acid
- Procedure: Mix 0.5 mL of PIA solution (appropriately diluted) with 0.5 mL of 5% phenol solution. Add 2.5 mL of concentrated sulfuric acid rapidly and vortex immediately. Allow to stand for 10 minutes, then vortex again. Incubate at room temperature for 20-30 minutes.
- Measurement: Read absorbance at 490 nm against a reagent blank.
- Standard Curve: Prepare using D-glucose or N-acetylglucosamine standards (0-100 μg/mL). Express results as glucose or GlcNAc equivalents [30].
N-Acetyl Group Quantification - Morgan-Elson Assay:
- Reagents: 0.8 M sodium borohydride, 0.5 M potassium tetraborate (pH 9.1), dimethylaminobenzaldehyde (DMAB) reagent (10 g DMAB in 100 mL of acetic acid containing 12.5% v/v 10 N HCl)
- Procedure: Mix 0.5 mL of PIA solution with 0.1 mL of sodium borohydride solution. Incubate at room temperature for 30 minutes. Add 0.1 mL of potassium tetraborate solution and heat at 100°C for 3 minutes. Cool, add 3 mL of DMAB reagent, and incubate at 37°C for 20 minutes.
- Measurement: Read absorbance at 585 nm.
- Standard Curve: Prepare using N-acetylglucosamine standards (0-100 μg/mL) [30].
These colorimetric methods enable researchers to quantify the total polysaccharide content and degree of acetylation, both critical parameters for understanding PIA's structure-function relationship in biofilms.
Data Presentation and Analysis
Spectral Signatures and Their Structural Correlations
Table 1: Key FTIR Absorption Bands for PIA Structural Analysis
| Wave Number (cm⁻¹) | Vibration Assignment | Structural Information | PIA-Specific Significance |
|---|---|---|---|
| 3300-3400 | N-H stretching | Amide A band | Indicates presence of N-acetyl groups; affected by deacetylation |
| 1650-1660 | C=O stretching | Amide I band | Confirms N-acetyl groups on GlcNAc residues |
| 1550-1560 | N-H bending, C-N stretching | Amide II band | Sensitivity to deacetylation; decreases with N-deacetylation |
| 1300-1450 | C-H bending | CH₂ and CH₃ deformations | Carbohydrate backbone characterization |
| 1030-1080 | C-OH stretching | Carbohydrate backbone | Confirms polysaccharide nature; sensitive to glycosidic linkages |
Table 2: Characteristic NMR Chemical Shifts for PIA/PNAG
| Nucleus | Chemical Shift (δ, ppm) | Assignment | Structural Significance |
|---|---|---|---|
| ( ^1H ) | 4.5-5.5 | Anomeric protons | Confirms β-configuration through coupling constants (J₁,₂ ≈ 8 Hz) |
| ( ^1H ) | 1.9-2.1 | N-acetyl methyl protons | Quantifies degree of acetylation; integral correlates with acetyl content |
| ( ^1H ) | 3.0-4.0 | Ring protons (H-2 to H-6) | Complex pattern confirms glucosamine backbone |
| ( ^{13}C ) | 100-105 | Anomeric carbons | Characteristic of β-(1→6) glycosidic linkages in PIA |
| ( ^{13}C ) | 174-176 | N-acetyl carbonyl carbons | Confirms presence of N-acetyl groups |
| ( ^{13}C ) | 22-25 | N-acetyl methyl carbons | Correlates with acetylation degree |
| ( ^{13}C ) | 50-60 | C-2 carbon (amine-bearing) | Shifts upon N-acetylation; sensitive to deacetylation |
Table 3: Colorimetric Assay Parameters for PIA Quantification
| Assay | Target Molecule/Group | Wavelength (nm) | Standard Compound | Application in PIA Analysis |
|---|---|---|---|---|
| Phenol-Sulfuric Acid | Total carbohydrates | 490 | D-glucose or GlcNAc | Quantifies total polysaccharide content in PIA preparations |
| Morgan-Elson | N-acetyl groups | 585 | N-acetylglucosamine | Determines degree of acetylation; critical for PIA functionality |
| Bradford | Protein contaminants | 595 | Bovine serum albumin | Assesses purity of PIA preparations; detects protein contamination |
Research Reagent Solutions for PIA Analysis
Table 4: Essential Research Reagents for PIA Structural Analysis
| Reagent/Material | Function/Application | Technical Specifications | Considerations for PIA Research |
|---|---|---|---|
| C18 Purification Column | Final purification of PIA | Reverse-phase chromatography | Removes residual lipids and hydrophobic contaminants after proteinase treatment |
| Proteinase K | Protein degradation | Microbial serine protease | Eliminates protein contaminants from PIA preparations; essential for pure samples |
| Centriprep 10 Device | Concentration and desalting | 10 kDa molecular weight cut-off | Retains PIA while removing small molecules and salts |
| Deuterated Solvents (D₂O, DMSO-d₆) | NMR spectroscopy | 99.8% deuterium or higher | Essential for NMR analysis; D₂O for native PIA, DMSO-d₆ for solubility challenges |
| Potassium Bromide (KBr) | FTIR sample preparation | FTIR grade, purity >99% | For preparing transparent pellets for transmission FTIR spectroscopy |
| N-acetylglucosamine | Standard for colorimetric assays | Analytical standard grade | Primary standard for Morgan-Elson assay and carbohydrate quantification |
| Dimethylaminobenzaldehyde (DMAB) | Morgan-Elson reagent | Colorimetric grade | Forms colored complex with N-acetylglucosamine derivatives |
Workflow Visualization
PIA Structural Analysis Workflow
Structural Features and Detection Methods
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), is a key constituent of the extracellular matrix responsible for biofilm formation in Staphylococcus aureus and Staphylococcus epidermidis [3] [1]. This cationic, partially deacetylated polymer of β-1-6-linked N-acetylglucosamine mediates bacterial cell-to-cell adhesion, facilitating the formation of structured microbial communities encased within a protective matrix [3] [35]. The clinical significance of PIA extends to its role in device-related infections, where it contributes to antibiotic resistance and immune evasion, making it a critical target for therapeutic intervention [3] [2]. This technical guide provides researchers with comprehensive methodologies for detecting and quantifying PIA-dependent biofilm formation in vitro, establishing crucial links between genetic determinants, molecular expression, and phenotypic output.
PIA Biosynthesis and Genetic Regulation
The icaADBC Operon and PIA Biosynthesis
PIA biosynthesis is mediated by the icaADBC operon (intercellular adhesion), which encodes enzymes responsible for the synthesis, export, and modification of the exopolysaccharide [3] [1]. The structural and functional relationships of these components are outlined below:
Figure 1: Genetic regulation and biosynthesis pathway of PIA in staphylococci
The icaADBC operon produces four essential proteins that function coordinately in PIA production [1] [35]. IcaA is an N-acetylglucosaminyltransferase that synthesizes PIA oligomers from UDP-N-acetylglucosamine, while IcaD forms a complex with IcaA to enhance its enzymatic efficiency [1]. IcaC is implicated in the externalization and elongation of the nascent polysaccharide chain, and IcaB functions as an N-deacetylase responsible for the partial deacetylation (approximately 15-20%) of PIA, conferring its positive charge essential for adhesive properties [3] [1] [35]. This deacetylation is crucial for the ability of PIA to mediate intercellular adhesion and confer resistance to antimicrobial peptides [35].
Environmental Regulation of PIA Production
PIA biosynthesis is strongly influenced by environmental conditions, which can modulate ica operon expression [3] [17]. Factors such as high salt concentrations, glucose availability, anaerobiosis, and iron limitation have been demonstrated to enhance PIA production and biofilm formation [17] [35]. This environmental regulation has significant implications for in vitro assay design, as standard laboratory growth conditions may not optimally induce PIA expression.
Essential Research Reagents and Materials
Table 1: Key research reagents for PIA-dependent biofilm studies
| Reagent Category | Specific Examples | Function in Biofilm Research |
|---|---|---|
| Growth Media | Tryptic Soy Broth (TSB), Brain Heart Infusion (BHI) | Base nutrient source for bacterial cultivation |
| Biofilm Inducers | Glucose (1%), Sucrose (2%), NaCl (4%) | Enhance PIA production and biofilm formation [18] [36] |
| Detection Reagents | Crystal violet (0.1-1%), Congo red | Stain biofilm biomass for quantification |
| Molecular Biology Tools | Primers for icaA, icaD genes; Proteinase K, DNase I | Confirm genetic basis of PIA production; analyze matrix composition [36] [31] |
| Enzymatic Inhibitors | Dispersin B (PNAG-degrading enzyme), Sodium metaperiodate | Specifically target PIA to confirm functional role [2] |
| Analytical Instruments | Microtiter plate reader (OD595/OD630), FTIR, NMR spectrometer | Quantify biofilm formation; characterize PIA structure [18] [31] |
Phenotypic Assays for PIA-Dependent Biofilm Detection
Tissue Culture Plate (TCP) Method: A Quantitative Approach
The Tissue Culture Plate (TCP) method represents the gold standard for quantitative assessment of biofilm formation, providing reliable, reproducible data suitable for statistical analysis [36].
Protocol:
- Inoculum Preparation: Adjust bacterial suspension in TSB to OD600 ≈ 0.7, then dilute 1:200 in appropriate growth medium [18]
- Media Supplementation: Enhance PIA production using:
- Incubation: Aliquot 200 µL diluted suspension into polystyrene microtiter plates, incubate statically for 24 hours at 37°C [18] [36]
- Washing: Remove planktonic cells by washing wells three times with phosphate-buffered saline (PBS)
- Fixation and Staining: Air-dry plates, stain with 150 µL of 1% crystal violet for 15 minutes [36]
- Destaining and Quantification: Elute bound stain with 30% acetic acid (160 µL), measure absorbance at 595 nm spectrophotometrically [18] [36]
Interpretation:
- OD595 < 0.120: Weak/non-biofilm producer
- OD595 0.120-0.240: Moderate biofilm producer
- OD595 > 0.240: Strong biofilm producer [36]
Congo Red Agar (CRA) Method: A Qualitative Screening Tool
The Congo Red Agar method provides a rapid, cost-effective approach for preliminary screening of biofilm-forming staphylococcal isolates [36].
Protocol:
- Medium Preparation: Prepare CRA containing 37 g/L brain heart infusion broth, 50 g/L sucrose, 10 g/L agar, and 0.8 g/L Congo red dye [36]
- Inoculation: Streak test isolates on CRA plates, incubate at 37°C for 24-48 hours
- Interpretation:
- Black, crystalline colonies: Strong biofilm producers
- Pink-red colonies: Non-biofilm producers [36]
While this method offers simplicity, it demonstrates lower sensitivity (14%) compared to genotypic methods, making it more suitable for initial screening rather than definitive characterization [36].
Tube Method (TM): A Semi-Quantitative Alternative
The Tube Method provides a simple approach for visualizing biofilm formation without specialized equipment [36].
Protocol:
- Inoculation: Incubate test strains in TSB with appropriate supplements (e.g., 1% glucose) in glass or polystyrene tubes for 18-24 hours at 37°C
- Visual Assessment: After gentle pouring, stain adherent film with crystal violet or directly observe
- Scoring:
- 0: No visible film
- 1: Weak film lining tube wall
- 2: Moderate film with ring formation
- 3: Strong, visible film [36]
This method demonstrates moderate sensitivity (64%) and specificity (74%) compared to genotypic detection of ica genes [36].
Method Validation and Data Interpretation
Comparative Analysis of Phenotypic Assays
Table 2: Performance characteristics of PIA biofilm detection methods
| Method | Sensitivity | Specificity | Advantages | Limitations |
|---|---|---|---|---|
| Tissue Culture Plate (with glucose/sucrose) | 59-80% [36] | 57% [36] | Quantitative, high-throughput, suitable for statistical analysis | Requires specialized equipment, more time-consuming |
| Congo Red Agar | 14% [36] | 86% [36] | Simple, cost-effective, no specialized equipment needed | Low sensitivity, qualitative results only |
| Tube Method | 64% [36] | 74% [36] | Simple visualization, semi-quantitative assessment | Subjective interpretation, limited quantification |
| Genotypic (icaAD detection) | Gold standard [36] | Gold standard [36] | Direct detection of genetic potential, highly specific | Does not confirm phenotypic expression, requires molecular biology facilities |
Establishing the PIA-Biofilm Link: Specificity Testing
To conclusively link biofilm formation to PIA production, researchers should implement specificity controls:
Enzymatic Disruption:
- Dispersin B: Specifically degrades PNAG/PIA; significant reduction in biofilm indicates PIA-dependent mechanism [2]
- Sodium metaperiodate: Targets polysaccharide components; sensitivity confirms exopolysaccharide-dependent biofilm
- Proteinase K: Digests protein-based adhesins; resistance suggests PIA predominance [31]
- DNase I: Degrades extracellular DNA; minimal effect indicates PIA-dependent biofilm [2]
Genetic Confirmation:
- PCR detection of icaA and icaD genes confirms genetic potential for PIA production [36]
- Isogenic mutant strains (e.g., ica-negative mutants) provide essential controls for PIA-specific phenotypes [18]
The experimental workflow for establishing this link is systematic:
Figure 2: Experimental workflow for establishing PIA-dependent biofilm formation
Advanced Applications and Research Implications
Therapeutic Applications: Anti-PIA Antibodies
Recent investigations have explored the potential of anti-PIA antibodies as therapeutic interventions. Immunization with purified PIA induces protective antibodies that demonstrate significant biofilm inhibition (approximately 40% opsonization properties) in both staphylococcal species and Escherichia coli, which produces a biochemically indistinguishable exopolysaccharide [18]. These antibodies effectively reduce biofilm formation in vitro, suggesting potential for immunoprophylactic approaches against device-related infections [18] [31].
In Vitro-In Vivo Correlation Challenges
Research comparing biofilm formation capacities of S. aureus strains Newman and Newman D2C revealed substantial differences in vitro that did not necessarily correlate with in vivo infectivity in foreign body-related murine infection models [17]. This highlights the critical importance of environmental context in PIA expression and function, suggesting that in vitro conditions must be carefully optimized to reflect clinically relevant scenarios.
Linking PIA to phenotypic output in vitro requires a multifaceted approach combining genetic confirmation with appropriate phenotypic assays under inducing conditions. The TCP method with glucose or sucrose supplementation provides the most reliable correlation with genotypic methods for detecting PIA-dependent biofilms [36]. Specificity should be confirmed through enzymatic disruption with PIA-targeting agents like Dispersin B, and researchers should remain cognizant of the potential discordance between in vitro biofilm formation capacity and in vivo pathogenicity. As PIA continues to be investigated as a vaccine target and therapeutic focus, robust and standardized in vitro assays will remain essential tools for advancing our understanding of staphylococcal biofilm pathogenesis and developing effective countermeasures.
Polysaccharide intercellular adhesin (PIA) has emerged as a principal virulence factor in staphylococcal biofilm formation and a promising target for vaccine development against device-related infections. Staphylococcus epidermidis and Staphylococcus aureus, the leading etiologic agents of implant-related infections, rely on PIA production for their ability to form adherent, multilayered biofilms on medical devices [2]. This exopolysaccharide, also known as poly-β(1,6)-N-acetylglucosamine (PNAG), forms a crucial component of the extracellular matrix that protects bacterial communities from host immune defenses and antibiotic treatments [3]. The clinical significance of PIA is underscored by epidemiological data showing that staphylococcal strains possessing the icaADBC biosynthetic locus are disproportionately associated with device-associated infections compared to colonizing isolates [3]. This review comprehensively examines PIA as a vaccine candidate, focusing on its structural characteristics, the functional properties of induced antibodies, experimental methodologies for evaluating immunogenicity, and recent advances in conjugate vaccine strategies aimed at enhancing opsonic and protective responses.
Structural and Functional Basis of PIA as a Vaccine Target
Biochemical Structure and Biosynthesis
PIA is a linear polymer of β-1,6-linked N-acetylglucosamine residues that are partially deacetylated (approximately 15-20%), conferring a positive charge to the molecule that is critical for its adhesive properties [37] [2]. The deacetylation process is mediated by IcaB, a key enzyme in the PIA biosynthetic pathway [11]. This cationic nature enables electrostatic interactions with negatively charged bacterial surfaces and host components, facilitating both cell-to-cell adhesion and immune evasion mechanisms [11].
The synthesis of PIA is governed by the icaADBC operon, which encodes the essential enzymes for polymer production and modification [3] [11]. Specifically, IcaA and IcaD generate the N-acetylglucosamine oligomers, IcaC facilitates externalization of the nascent polysaccharide, and IcaB performs the crucial deacetylation step that enables surface attachment and intercellular adhesion [2] [11]. The regulatory control of this operon is complex, responding to environmental conditions through various regulatory systems, and subject to phase variation that creates heterogeneous expression patterns within bacterial populations [3] [2].
Role in Biofilm Formation and Immune Evasion
PIA contributes significantly to staphylococcal pathogenicity through dual mechanisms of biofilm structural integrity and immune system evasion. As the primary mediator of cell-to-cell adhesion in many staphylococcal strains, PIA enables the formation of stable, three-dimensional biofilm architectures that resist mechanical disruption [37]. Beyond this structural role, PIA provides protection against innate host defenses by inhibiting neutrophil phagocytosis and complement activation, allowing biofilm-embedded bacteria to persist in hostile host environments [18] [3]. This functional combination makes PIA an ideal vaccine target, as antibodies disrupting these mechanisms could potentially mitigate both biofilm formation and immune resistance.
Table 1: Key Structural and Functional Characteristics of PIA
| Characteristic | Description | Functional Significance |
|---|---|---|
| Chemical Structure | Linear polymer of β-1,6-linked N-acetylglucosamine residues | Forms stable extracellular matrix |
| Deacetylation | 15-20% of residues non-N-acetylated | Confers positive charge for surface attachment and immune evasion |
| Molecular Weight | Variable reports (~30 kDa to >460 kDa) | Size variations may affect antigenic properties |
| Genetic Basis | Encoded by icaADBC operon | Conservation across staphylococcal species and some Gram-negative pathogens |
| Biosynthetic Regulation | Influenced by environmental conditions and multiple regulatory systems | Affects expression heterogeneity and vaccine target consistency |
Experimental Models for Evaluating PIA Vaccine Efficacy
PIA Extraction and Purification Protocols
The standardized methodology for PIA extraction begins with culturing S. epidermidis in tryptic soy broth under mild agitation conditions (40-50 rpm/min) for 24 hours at 37°C [18] [30]. Bacterial cells are harvested via centrifugation at 1000 G for 20 minutes at 4°C, followed by resuspension in phosphate-buffered saline (PBS, pH 7.5) and sonication on ice (four cycles of 30 seconds each) [30]. After further centrifugation at 12,000 rpm for 15 minutes, the supernatant undergoes concentration using Centriprep 10 devices (Amicon, Germany) [30] [31]. Subsequent purification steps include enzymatic digestion with proteinase K to eliminate soluble proteins, followed by chromatography using C18 columns or Sephacryl S-100 matrix equilibrated with 50 mM PBS [18] [31]. The structural confirmation of purified PIA is essential and is typically achieved through Fourier Transform Infrared Spectroscopy (FTIR), Nuclear Magnetic Resonance (NMR), and colorimetric assays targeting hexosamine content [18] [31].
Animal Immunization Strategies
Recent investigations have employed mouse models to evaluate the immunogenicity of PIA-based vaccines. A standard protocol involves immunizing female BALB/c mice (6-8 weeks old) subcutaneously on days 0, 14, and 28 with 100 μg of PIA antigen formulated with 1% alum adjuvant [31]. Blood collection via the orbital sinus two weeks after each immunization enables serial assessment of antibody titers. Studies consistently demonstrate that this immunization schedule elicits significant increases in anti-PIA IgG levels compared to control groups, with particularly robust boosts following secondary immunization [18] [31]. For conjugate vaccines, such as PIA-rSesC, similar immunization schedules have proven effective in generating opsonic antibodies that protect against challenge with wild-type S. epidermidis [38].
Analytical Techniques for Immune Response Characterization
The evaluation of humoral responses to PIA vaccination employs multiple complementary techniques. Enzyme-linked immunosorbent assay (ELISA) represents the cornerstone method for quantifying anti-PIA antibody titers, typically using 1 μg/well of purified PIA for coating and serial dilutions of immune sera [31]. Beyond total IgG measurement, subclass analysis (particularly IgG2a and IgG2b) provides insights into the Th1/Th2 polarization of the immune response [38]. Flow cytometry protocols, following established immunological guidelines, enable detailed characterization of immune cell populations and surface marker expression in response to vaccination [39]. For functional assessment, opsonophagocytic killing assays quantify the ability of anti-PIA antibodies to promote bacterial clearance by immune cells, with recent studies reporting approximately 40.48% opsonization-mediated lethality against E. coli producing the homologous PNAG polysaccharide [18] [30].
Quantitative Assessment of PIA Vaccine Efficacy
Antibody-Mediated Biofilm Inhibition
The capacity of PIA-specific antibodies to disrupt biofilm formation has been quantitatively demonstrated through standardized microtiter plate assays. Immunized mouse sera containing anti-PIA antibodies exhibit substantial biofilm inhibitory effects compared to non-immune sera [18] [31]. The percentage of biofilm inhibition is calculated using the formula: [(A595, positive - A595, antibody)/(A595, positive - A595, negative)] × 100, where A595 represents absorbance at 595nm after crystal violet staining [31]. Studies implementing this methodology have confirmed that PIA-induced antibodies significantly impair biofilm formation in both S. epidermidis and S. aureus wild-type strains [31]. Notably, the inhibitory efficacy appears enhanced when PIA is administered in combination with other staphylococcal antigens, such as glycerol teichoic acid (Gly-TA), suggesting potential synergistic effects in multi-component vaccine formulations [31].
Cross-Species Protective Efficacy
Remarkably, PIA-induced antibodies demonstrate cross-protective activity against bacterial species producing structurally similar exopolysaccharides. Recent research reveals that anti-PIA antibodies generated through immunization with staphylococcal PIA exhibit 40.48% opsonic killing efficacy against Escherichia coli ATCC 25922, which produces PNAG biochemically indistinguishable from PIA [18] [30]. This cross-reactivity stems from the structural conservation of the poly-β(1,6)-N-acetylglucosamine backbone across taxonomically diverse bacterial pathogens, including Yersinia pestis, Bordetella species, Klebsiella pneumoniae, and Acinetobacter baumannii [3]. The functional implications of this cross-reactivity are significant, suggesting that PIA-based vaccines could potentially provide broad protection against multiple biofilm-forming pathogens beyond staphylococci.
Table 2: Quantitative Efficacy Measures of PIA-Based Vaccines from Recent Studies
| Vaccine Formulation | Immunization Model | Antibody Response | Protective Efficacy | Reference |
|---|---|---|---|---|
| Purified PIA | Mouse model (3 immunizations) | Significant increase in IgG titers (p=0.0021) | 40.48% opsonic killing vs E. coli; Significant biofilm inhibition | [18] [30] [31] |
| PIA-rSesC Conjugate | Mouse model | Elevated opsonic antibody levels | Protection against S. epidermidis challenge; Biofilm elimination in vitro and in vivo | [38] |
| PIA/Gly-TA Mixture | Mouse model (50/50 μg) | Enhanced IgG response post-booster (p=0.0123) | Superior biofilm inhibition vs single antigens | [31] |
Conjugate Vaccine Strategies to Enhance PIA Immunogenicity
Rationale for Conjugate Approaches
As a pure polysaccharide antigen, PIA typically elicits T-cell-independent immune responses characterized by limited immunoglobulin class switching and poor immunological memory. Conjugate vaccine technology, which links polysaccharide antigens to protein carriers, overcomes this limitation by engaging T-cell help and promoting robust, long-lasting immunity [38]. The protein component of conjugate vaccines facilitates antigen processing and presentation via MHC class II molecules, thereby activating helper T cells and enabling the development of memory B cell populations. This approach has proven highly successful in vaccine development against other polysaccharide-encapsulated pathogens, including Haemophilus influenzae type b and Streptococcus pneumoniae.
Implementation and Efficacy of PIA Conjugates
Recent investigations have explored the conjugation of PIA to various protein carriers to enhance immunogenicity. One promising approach involves covalent linkage of PIA to recombinant SesC (rSesC), a truncated staphylococcal surface protein [38]. Structural analysis of the resulting PIA-rSesC conjugate via FTIR and 1H-NMR confirms successful conjugation while maintaining the structural integrity of both components [38]. Immunization with this conjugate vaccine induces significantly higher levels of opsonic antibodies compared to PIA alone, with demonstrated efficacy in protecting mice against intravenous challenge with wild-type S. epidermidis [38]. Importantly, vaccinated mice show substantial reduction in bacterial biofilm formation in both in vitro and in vivo assays, validating the functional relevance of the enhanced immune response [38].
Research Reagent Solutions for PIA Vaccine Development
Table 3: Essential Research Reagents for PIA Vaccine Evaluation
| Reagent/Category | Specific Examples | Research Application | Functional Role |
|---|---|---|---|
| Analytical Instruments | FTIR Spectrometer, NMR | Structural confirmation of purified PIA | Verification of polysaccharide structure and conjugation efficacy |
| Immunoassays | ELISA kits, ELISpot/FluoroSpot assays | Quantification of antibody titers and antigen-specific B cells | Immune response monitoring and cellular immunity characterization |
| Chromatography Systems | C18 columns, Sephacryl S-100 matrix | PIA purification and conjugate separation | Isolation of high-purity antigen for immunization |
| Cell Culture Components | Tryptic Soy Broth, Mueller Hinton Agar | Bacterial culture and antibiotic susceptibility testing | Strain maintenance and phenotypic confirmation |
| Animal Model Supplies | BALB/c mice, alum adjuvant | In vivo immunogenicity and protection studies | Preclinical vaccine efficacy evaluation |
| Biofilm Assay Materials | Polystyrene microtiter plates, crystal violet | Quantitative assessment of biofilm inhibition | Functional evaluation of antibody efficacy |
PIA represents a compelling vaccine candidate against staphylococcal biofilm-associated infections, with demonstrated capacity to elicit opsonic and protective antibodies in preclinical models. The structural conservation of this poly-β(1,6)-N-acetylglucosamine polymer across diverse bacterial pathogens further enhances its attractiveness as a broad-spectrum target. Current evidence confirms that PIA-specific antibodies significantly inhibit biofilm formation and promote bacterial clearance through opsonophagocytosis. Conjugate vaccine strategies markedly enhance PIA immunogenicity by engaging T-cell-dependent immune mechanisms, resulting in more robust and durable protective responses. Future research directions should include optimization of conjugation methods, exploration of novel adjuvant systems, expansion of cross-protection studies against additional PIA-producing pathogens, and progression toward clinical evaluation in human populations. The continued development of PIA-based vaccines holds substantial promise for addressing the pressing clinical challenge of device-related infections, particularly in an era of increasing antibiotic resistance.
Biofilm-associated infections represent a significant challenge in clinical settings, accounting for 65-80% of all human microbial infections [40]. Within the context of Staphylococcus biofilm research, the polysaccharide intercellular adhesin (PIA) has emerged as a fundamental virulence determinant enabling persistent infections on medical implants and devices [28] [19]. PIA is a poly-β(1-6)-N-acetylglucosamine (PNAG) that is partially deacetylated, giving it a positive charge that facilitates intercellular adhesion and structural integrity of the staphylococcal biofilm matrix [28]. The synthesis of PIA is mediated by the icaADBC operon, with IcaA functioning as an N-acetylglucosaminyltransferase that synthesizes PIA oligomers, IcaD enhancing IcaA efficiency, IcaC involved in polysaccharide externalization, and IcaB serving as an N-deacetylase responsible for the partial deacetylation of PIA [28]. This extracellular polymeric substance (EPS) forms a protective shield for encased bacterial cells, providing remarkable resistance to antibiotics and host immune responses [40].
The significance of PIA in Staphylococcal biofilms is particularly evident in prosthetic joint infections (PJI), where Staphylococcus aureus and Staphylococcus epidermidis are the most frequently isolated pathogens [19]. Research has demonstrated that biofilm formation is a common phenotypic feature in staphylococci isolated from PJI, mediated by both PIA and protein factors [19]. Interestingly, these different factors appear to have differential importance in the pathogenesis of S. epidermidis PJI, with PIA being particularly significant in total knee arthroplasty (TKA) related infections, while in total hip arthroplasty (THA) protein factors like the accumulation-associated protein (Aap) are sufficient to establish persistent infections [19]. In S. aureus, both PIA and surface proteins like SasG work in parallel to mediate biofilm accumulation [19].
Against this backdrop, enzymatic disruption strategies targeting key biofilm matrix components have emerged as promising therapeutic approaches. Dispersin B and DNase I represent two distinct enzymatic strategies that specifically target the polysaccharide (PIA) and extracellular DNA (eDNA) components of the biofilm matrix, respectively [28] [41]. This technical review examines the mechanisms, efficacy, and application protocols for these enzymatic anti-biofilm agents within the framework of PIA-centric staphylococcal biofilm research.
Biofilm Matrix Composition: PIA as a Therapeutic Target
The extracellular polymeric substance (EPS) of bacterial biofilms is a complex matrix consisting of extracellular proteins, lipids, nucleic acids, polysaccharides, and secondary metabolites [40]. In staphylococcal biofilms, PIA serves as a primary structural scaffold, with its partially deacetylated poly-β(1-6)-N-acetylglucosamine structure facilitating cation-mediated intercellular adhesion and matrix stability [28]. The protective function of this matrix is twofold: it acts as a physical barrier against exogenous stress and creates heterogeneous microenvironments where bacterial cells exhibit reduced metabolic rates and increased antibiotic tolerance [40].
Notably, the biofilm matrix composition varies among methicillin-resistant Staphylococcus aureus (MRSA) clones, with transcriptional studies revealing striking differences in gene expression during biofilm formation among pandemic clones like USA300, HEMRSA-15, and ST239 [42]. While the icaADBC locus responsible for PIA synthesis is widespread in clinically significant staphylococcal isolates, MRSA has been shown to form more proteinaceous biofilms independent of the icaADBC mechanism that prevails in methicillin-sensitive S. aureus (MSSA) biofilms [42]. This highlights the importance of combination therapies targeting multiple matrix components.
Table 1: Major Components of the Staphylococcal Biofilm Matrix and Their Functions
| Matrix Component | Chemical Structure | Primary Function | Genetic Regulation |
|---|---|---|---|
| Polysaccharide Intercellular Adhesin (PIA) | Poly-β(1-6)-N-acetylglucosamine (partially deacetylated) | Intercellular adhesion, structural scaffold, cation chelation | icaADBC operon |
| Extracellular DNA (eDNA) | Double-stranded DNA fragments | Structural integrity, cation chelation, horizontal gene transfer | Autolysis-regulated release |
| Biofilm-associated proteins (Bap) | Amyloid-like proteins with multiple repeats | Surface adhesion, structural stability | bap gene |
| Accumulation-associated protein (Aap) | Membrane-bound protein | PIA-independent intercellular adhesion | aap gene |
| Extracellular RNA | Various RNA species | Structural stability, genetic regulation | Poorly characterized |
The following diagram illustrates the key stages in the development of a PIA-dependent staphylococcal biofilm and the strategic intervention points for enzymatic disruption:
Diagram 1: Staphylococcal Biofilm Lifecycle and Enzymatic Intervention Points. The diagram illustrates the key developmental stages of PIA-dependent biofilms and the strategic targeting by Dispersin B (glycoside hydrolase) and DNase I (nuclease) at specific maturation phases.
Enzymatic Mechanisms of Action
Dispersin B: Glycoside Hydrolase Targeting PIA/PNAG
Dispersin B is a glycoside hydrolase that specifically targets the poly-β(1,6)-N-acetyl-d-glucosamine (PNAG) polysaccharide, which constitutes the structural backbone of PIA in staphylococcal biofilms [40] [28]. This enzyme catalyzes the hydrolysis of the β(1,6) glycosidic linkages in the PNAG polymer, effectively depolymerizing the exopolysaccharide scaffold and disrupting the intercellular adhesion between bacterial cells [40]. The enzymatic degradation of PIA leads to loss of biofilm integrity, detachment of sessile cells, and conversion to the more susceptible planktonic phenotype.
The specificity of Dispersin B for PNAG is particularly relevant for targeting staphylococcal biofilms, as dPNAG has been identified as a key exopolysaccharide in many medically relevant pathogens including Staphylococcus aureus, Staphylococcus epidermidis, and numerous other Gram-positive and Gram-negative bacteria [40]. The enzyme functions extracellularly without needing to cross bacterial membranes, making resistance development less likely compared to conventional antibiotics [40]. Laboratory studies have demonstrated that Dispersin B can be procured at high concentrations through isolation or over-expression in model organisms and added exogenously to microbial colonies to efficiently break down pre-existing biofilms [40].
DNase I: Nuclease Targeting Extracellular DNA (eDNA)
DNase I represents a complementary enzymatic strategy that targets the extracellular DNA (eDNA) component of the biofilm matrix [28] [41]. eDNA serves as a critical structural element in many bacterial biofilms, providing structural integrity through cation chelation and functioning as a cell-to-cell adhesion molecule [41]. In staphylococcal biofilms, eDNA works in concert with PIA to stabilize the three-dimensional architecture of the matrix, with the relative importance of each component varying between strains and environmental conditions.
DNase I cleaves eDNA through endonucleolytic activity, generating oligonucleotides with 5'-phospho and 3'-hydroxyl termini [41]. This degradation of the eDNA framework disrupts matrix stability and facilitates biofilm disintegration. The efficacy of DNase I against staphylococcal biofilms is particularly notable in MRSA strains that produce proteinaceous or eDNA-rich biofilms, as well as in polymicrobial biofilms where eDNA may represent a more conserved structural component across species [41].
Table 2: Comparative Analysis of Dispersin B and DNase I Anti-Biofilm Enzymes
| Characteristic | Dispersin B | DNase I |
|---|---|---|
| Enzyme Class | Glycoside hydrolase | Nuclease |
| EC Classification | EC 3.2.1.- | EC 3.1.21.1 |
| Primary Target | β(1,6)-N-acetyl-D-glucosamine polymer (PNAG/PIA) | Double-stranded DNA |
| Mechanism of Action | Hydrolysis of glycosidic bonds in polysaccharide backbone | Endonucleolytic cleavage of phosphodiester bonds |
| Optimal pH Range | 5.5-6.5 | 7.0-8.0 |
| Cofactor Requirements | None | Ca²⁺ and Mg²⁺ or Mn²⁺ |
| Biofilm Reduction Efficacy | 70-90% against PIA-dependent staphylococcal biofilms | 60-85% against eDNA-rich biofilms |
| Effective Concentration Range | 10-100 μg/mL | 5-50 μg/mL |
| Synergy with Antibiotics | Enhances efficacy of aminoglycosides, β-lactams, glycopeptides | Enhances efficacy of fluoroquinolones, macrolides |
Experimental Protocols for Enzymatic Biofilm Disruption
Standardized Biofilm Cultivation Model
For consistent evaluation of anti-biofilm efficacy, researchers should employ standardized biofilm cultivation techniques. The static biofilm model using high-nutrient media supplemented with conditioning agents provides reproducible results for staphylococcal biofilms:
Inoculum Preparation: Grow test strains (e.g., S. aureus ATCC 6538, S. epidermidis ATCC 35984) in brain heart infusion (BHI) broth supplemented with 0.1-1% glucose at 37°C with shaking (150-200 rpm) to mid-exponential phase (OD₆₀₀ ≈ 0.5-0.8) [42].
Biofilm Formation: Dilute cultures to approximately 1×10⁶ CFU/mL in fresh BHI with 1% glucose. Aliquot 100-200 μL per well into 96-well polystyrene microtiter plates. Include negative controls (uninoculated media) and positive controls (established biofilm formers) [43].
Incubation Conditions: Incubate plates statically at 37°C for 24-48 hours to allow biofilm development. For mature biofilms, extend incubation to 72-96 hours with media refreshment at 24-hour intervals [42].
Biofilm Quantification: Wash established biofilms gently with phosphate-buffered saline (PBS) to remove non-adherent cells. Fix with 99% methanol for 15 minutes, then stain with 0.1% crystal violet for 15-20 minutes. Wash excess stain and elute bound dye with 33% glacial acetic acid. Measure absorbance at 570-600 nm [43].
Enzymatic Treatment and Assessment
The following protocol details the application of Dispersin B and DNase I for biofilm disruption studies:
Enzyme Preparation:
- Prepare Dispersin B stock solution (1-2 mg/mL) in sterile 20 mM sodium phosphate buffer (pH 5.8-6.0) [28].
- Prepare DNase I stock solution (1 mg/mL) in sterile Tris-HCl buffer (pH 7.5-7.6) containing 1-2 mM CaCl₂ and MgCl₂ [41].
- Create working concentrations through serial dilution in appropriate buffers.
Enzymatic Treatment:
- Gently wash established 24-hour biofilms with PBS or appropriate buffer.
- Apply enzyme treatments (100 μL/well) at concentrations ranging from 1-100 μg/mL for Dispersin B and 1-50 μg/mL for DNase I.
- Include buffer-only controls and combination treatments for synergy studies.
- Incubate at 37°C for 1-24 hours depending on experimental endpoint.
Efficacy Assessment:
- Biomass Reduction: Quantify remaining biofilm using crystal violet staining as described above [43].
- Viability Assessment: Use metabolic assays (MTT/XTT) or colony counting after biofilm disruption to determine bactericidal effects.
- Morphological Analysis: Employ scanning electron microscopy (SEM) or confocal laser scanning microscopy (CLSM) to visualize structural changes [43].
- Synergy Testing: Combine sub-effective concentrations of enzymes with conventional antibiotics to determine fractional inhibitory concentration indices.
The following workflow diagram outlines the key procedural stages for conducting enzymatic biofilm disruption experiments:
Diagram 2: Experimental Workflow for Enzymatic Biofilm Disruption Studies. The diagram outlines the key procedural stages from biofilm cultivation through enzymatic treatment to efficacy assessment and synergy testing.
Research Reagent Solutions for Anti-Biofilm Studies
Table 3: Essential Research Reagents for Enzymatic Anti-Biofilm Studies
| Reagent/Category | Specifications & Functions | Application Notes |
|---|---|---|
| Dispersin B | Glycoside hydrolase (≥95% purity), specific for β-1,6-N-acetyl-D-glucosamine | Use concentration: 10-100 μg/mL; Buffer: Sodium phosphate (pH 5.8-6.0); Storage: -20°C |
| DNase I | Nuclease (≥85% purity), requires Ca²⁺/Mg²⁺ cofactors | Use concentration: 5-50 μg/mL; Buffer: Tris-HCl with Ca²⁺/Mg²⁺ (pH 7.5); Storage: -20°C |
| Biofilm Cultivation Media | Brain Heart Infusion (BHI) with 0.1-1% glucose | Enhanced EPS production; Filter-sterilize glucose separately |
| Microtiter Plates | 96-well, polystyrene, flat-bottom, tissue culture-treated | Standardized surface for biofilm formation; minimal lot-to-lot variation |
| Crystal Violet Stain | 0.1% aqueous solution, filtered | Biomass quantification; consistent staining time (15-20 min) |
| Metabolic Activity Assays | MTT (0.5 mg/mL) or XTT-menadione | Viability assessment; protect from light; fresh preparation |
| Confocal Microscopy Supplies | LIVE/DEAD BacLight Bacterial Viability Kits | 3D biofilm visualization; SYTO9 (5 μM) and propidium iodide (30 μM) |
| Antibiotic Panels | Clinically relevant classes: aminoglycosides, β-lactams, glycopeptides | Synergy studies; prepare fresh from powder or frozen stocks |
Comparative Efficacy and Synergistic Applications
Quantitative assessment of enzymatic anti-biofilm agents reveals distinct efficacy profiles against staphylococcal biofilms. Dispersin B demonstrates superior activity against PIA-dependent biofilms, typically achieving 70-90% biomass reduction at concentrations of 10-100 μg/mL within 2-4 hours of treatment [28]. The enzyme exhibits optimal activity at slightly acidic pH (5.5-6.5), which may enhance efficacy in certain infection environments like chronic wounds where the tissue pH is often acidic. DNase I shows broad-spectrum activity against diverse biofilm types, with particular effectiveness against eDNA-rich matrices (60-85% disruption at 5-50 μg/mL), and functions optimally at neutral to slightly alkaline pH (7.0-8.0) [41].
The strategic combination of these enzymes with conventional antibiotics represents a promising approach to combat biofilm-associated infections. Research demonstrates that enzymatic disruption of the biofilm matrix significantly enhances antibiotic penetration and efficacy against the embedded bacterial populations [40] [41]. Dispersin B pre-treatment has been shown to reduce the minimum biofilm eradication concentration (MBEC) of various antibiotics by 10-1000 fold, with particularly notable synergy observed with aminoglycosides, β-lactams, and glycopeptides against staphylococcal biofilms [40]. Similarly, DNase I enhances the efficacy of fluoroquinolones and macrolides, especially against MRSA strains that produce eDNA-rich biofilms [41].
The following diagram illustrates the synergistic mechanism of combined enzymatic and antibiotic treatment:
Diagram 3: Synergistic Mechanism of Combined Enzymatic and Antibiotic Therapy. The diagram illustrates how sequential application of matrix-degrading enzymes (Dispersin B and DNase I) followed by conventional antibiotics enhances biofilm eradication through improved antimicrobial penetration.
Transcriptional analyses of MRSA biofilms during enzymatic disruption reveal complex adaptive responses. Studies of pandemic MRSA clones (USA300, HEMRSA-15, ST239) demonstrate upregulation of genes involved in transporter systems and core genome-encoded KEGG pathways during early biofilm development [42]. The clumping factor A gene (clfA) emerges as a consistently upregulated factor across MRSA clones in 24-hour biofilms (2.2- to 2.66-fold increase), representing a potential secondary target for anti-biofilm strategies [42]. Enzymatic treatment triggers stress response pathways and alters expression of virulence regulators, highlighting the importance of targeting both structural and genetic determinants of biofilm formation.
Enzymatic disruption of staphylococcal biofilms using Dispersin B and DNase I represents a promising therapeutic strategy that specifically targets the structural integrity of the biofilm matrix. Within the context of PIA-focused staphylococcal research, these enzymes offer mechanistically distinct approaches to combat biofilm-mediated antibiotic resistance and treatment failure. The specificity of Dispersin B for PNAG/PIA makes it particularly valuable against PIA-dependent staphylococcal biofilms prevalent in medical device-related infections, while DNase I provides complementary activity against eDNA-rich matrices.
Future research directions should focus on optimizing delivery systems for these enzymes, exploring engineered variants with enhanced stability and activity, and conducting comprehensive in vivo efficacy studies. Additionally, investigation of enzymatic combinations with unconventional antimicrobial agents (e.g., antimicrobial peptides, phage therapy) may yield novel synergistic approaches. As our understanding of biofilm heterogeneity and genetic regulation advances, particularly through transcriptional studies of diverse MRSA clones [42], targeted enzymatic strategies will continue to evolve as essential components in the therapeutic arsenal against persistent biofilm-associated infections.
Beyond PIA: Alternative Mechanisms and Biofilm Heterogeneity
The formation of biofilms is a central virulence mechanism for staphylococci, enabling these pathogens to cause persistent infections, particularly on indwelling medical devices. For decades, research on Staphylococcus aureus and Staphylococcus epidermidis biofilms was dominated by the study of the polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG). PIA is a cationic, partially deacetylated homopolymer of β-1,6-linked N-acetylglucosamine, whose biosynthesis is directed by the icaADBC operon [3] [28]. While undoubtedly important, the singular focus on PIA obscured a critical reality: many clinical isolates form robust biofilms through PIA-independent mechanisms [44]. It is now established that a multitude of surface proteins can promote the accumulation phase of biofilm formation independently of the ica locus [45]. This whitepaper details the molecular pathways of ica-independent biofilm accumulation, focusing on the key proteinaceous factors Aap (accumulation-associated protein) and SasG (S. aureus surface protein G), and frames their significance within the broader context of staphylococcal biofilm research and therapeutic development.
Key Proteinaceous Mediators of Ica-Independent Biofilm Accumulation
The biofilm matrix in staphylococci is a complex mixture of macromolecules. In the absence of PIA, surface proteins can fulfill an analogous structural role, mediating both attachment to surfaces and critical intercellular interactions that underpin biofilm accumulation [45]. The following table summarizes the primary protein factors involved.
Table 1: Key Proteinaceous Factors in Ica-Independent Staphylococcal Biofilm Formation
| Protein Factor | Organism | Primary Domain(s) | Proposed Mechanism in Biofilm Accumulation | Regulatory Cofactor |
|---|---|---|---|---|
| Aap (Accumulation-Associated Protein) | S. epidermidis | A domain (L-type lectin), G5, E repeats | Zn²⁺-dependent homophilic interactions between B-repeat fibrils; requires A-domain proteolytic cleavage [45] [46]. | Zn²⁺ |
| SasG (S. aureus Surface Protein G) | S. aureus | A domain, G5, E repeats | Zn²⁺-dependent homophilic interactions between B-repeat domains; spontaneous cleavage may facilitate interaction [45] [46]. | Zn²⁺ |
| Bap (Biofilm-Associated Protein) | S. aureus (bovine) | Multiple repeat domains | Promotes intercellular adhesion and biofilm accumulation; mechanism distinct from SasG/Aap [45]. | Not Specified |
| FnBPA/FnBPB (Fibronectin-Binding Proteins A/B) | S. aureus | MSCRAMM family | Promote biofilm accumulation in some MRSA strains, potentially by bridging cells via fibronectin [45] [46]. | Not Specified |
| Emp (Extracellular Matrix Protein-binding protein) | S. aureus | Not Specified | Multifunctional protein that promotes biofilm formation under iron-depleted conditions [45]. | Fe²⁺ (depletion) |
Molecular Architecture and Mechanism of Aap and SasG
Structural Organization
Aap and its S. aureus ortholog SasG are structurally homologous LPXTG-anchored, cell wall-associated proteins [45] [46]. Their domain architecture is highly conserved, comprising several key regions: The N-terminal A domain contains a legume-like (L-type) lectin domain potentially involved in initial surface attachment [45]. Linking the A domain to the cell wall is the B region, which consists of a variable number of tandem repeats. Each repeat is made up of a G5 domain separated by a 50-residue E region [45] [46]. The proteins are anchored to the cell wall peptidoglycan via a C-terminal LPXTG motif recognized by sortase A [45].
The Zn²⁺-Dependent "Zipper" Model for Intercellular Adhesion
The current model for Aap- and SasG-mediated biofilm accumulation involves a specific, Zn²⁺-dependent homophilic interaction between B regions on adjacent bacterial cells [46].
Diagram: Molecular Mechanism of Aap/SasG-Mediated Biofilm Accumulation
The diagram above illustrates the core mechanism:
- Proteolytic Activation: For Aap, and potentially SasG, the N-terminal A domain is removed via proteolytic cleavage by bacterial or host proteases. This cleavage exposes the B-repeat region, which is necessary for its intercellular function [45] [46].
- Dimerization and Fibril Formation: The exposed B repeats, comprised of G5 and E domains, form extended fibrillar structures on the cell surface.
- Zn²⁺-Dependent Homophilic Interaction: Dimerization between G5-E fibrils from adjacent cells occurs in a Zn²⁺-dependent manner. The Zn²⁺ ion is chelated by histidine residues within the E region, forming a stable cross-bridge that "zips" the cells together [46]. This interaction is both specific and strong, providing the mechanical foundation for a proteinaceous biofilm matrix.
Experimental Characterization of Proteinaceous Biofilms: Key Methodologies
Establishing Protein-Dependent Biofilm Formation
The first step in characterizing an ica-independent biofilm is to confirm the proteinaceous nature of the matrix. This is typically achieved through enzymatic treatment of pre-formed biofilms [45].
- Protocol: Established biofilms, grown in microtiter plates or on other surfaces, are incubated with a protease such as trypsin or proteinase K (e.g., 1 mg/mL in appropriate buffer for 1-2 hours at 37°C). A parallel control biofilm is incubated with buffer alone.
- Interpretation: A significant reduction in biofilm integrity or biomass following protease treatment, but not in the control, is a clear indication of a protein-dependent matrix. This can be coupled with periodate treatment, which oxidizes polysaccharides, to rule out the involvement of PIA or other exopolysaccharides [45].
Genetic Validation via Site-Specific Mutagenesis
The contribution of a specific protein is definitively tested by creating isogenic mutant strains.
- Protocol: The gene encoding the protein of interest (e.g., aap or sasG) is inactivated via allelic replacement using an antibiotic resistance cassette. The biofilm-forming capacity of the mutant strain is then compared to the wild-type parent strain and a genetically complemented mutant in standard assays [45] [46].
- Advanced Genetic Tests: Loss of biofilm in a mutant defective in sortase A (the enzyme that anchors LPXTG proteins to the cell wall) suggests the involvement of one or more CWA proteins. Systematic inactivation of individual CWA protein genes can then pinpoint the specific factor responsible [45].
In Vitro Biofilm Assay (Microtiter Plate)
This is a foundational, high-throughput method for quantifying biofilm formation [20] [46].
- Protocol:
- Inoculation: Bacterial cultures are grown overnight, diluted (typically 1:200) in a rich medium like Tryptic Soy Broth (TSB) or Brain Heart Infusion (BHI), often supplemented with 1% glucose (BHIGlc).
- Growth: The diluted culture is aliquoted into the wells of a polystyrene microtiter plate (e.g., 200 µL per well) and incubated statically for 18-24 hours at 37°C.
- Staining: Planktonic cells are removed by washing with phosphate-buffered saline (PBS). Adherent cells are then fixed and stained with a 0.1% crystal violet solution for 15-30 minutes.
- Quantification: After washing off excess stain, the bound crystal violet is solubilized with a solution like 30% acetic acid or ethanol-acetone. The absorbance of the solution is measured spectrophotometrically at 570-595 nm, providing a quantitative measure of biofilm biomass [46].
Investigating the Role of Zn²⁺
The specific dependence on zinc can be tested using chelators and purified protein components [46].
- Protocol:
- In vivo: Biofilm assays are performed in media supplemented with the specific Zn²⁺ chelator diethylenetriaminepentaacetic acid (DTPA) (e.g., 100-500 µM). Inhibition of biofilm formation implicates a Zn²⁺-dependent mechanism.
- In vitro: Recombinant B-repeat proteins are purified and analyzed for Zn²⁺-dependent dimerization using techniques like size-exclusion chromatography or analytical ultracentrifugation in the presence and absence of Zn²⁺ and DTPA [46].
- Inhibition Assay: Purified recombinant B-domain protein can be added to growing bacterial cultures. This soluble protein can bind to surface-exposed B domains, blocking homophilic interactions and inhibiting biofilm formation in a dose-dependent manner [46].
The Scientist's Toolkit: Essential Research Reagents
The following table catalogues critical reagents used in the experimental analysis of Aap, SasG, and related proteinaceous biofilm factors.
Table 2: Research Reagent Solutions for Studying Proteinaceous Biofilms
| Reagent / Material | Function / Application | Example in Context |
|---|---|---|
| Proteinase K / Trypsin | Protease for degrading proteinaceous biofilm matrix; confirms protein-dependence. | Incubation with 1 mg/mL proteinase K disrupts Aap-dependent biofilms [45]. |
| Crystal Violet | Dye for staining and quantifying adherent biofilm biomass in microtiter assays. | Standard staining for high-throughput screening of biofilm-forming isolates [46] [44]. |
| Diethylenetriaminepentaacetic Acid (DTPA) | Specific chelator of Zn²⁺ ions; inhibits Zn²⁺-dependent homophilic interactions. | 100-500 µM DTPA in growth medium inhibits SasG-mediated biofilm accumulation [46]. |
| Polystyrene Microtiter Plates | Surface for high-throughput, in vitro biofilm formation assays. | Used in standard static biofilm assays to phenotype strains [20] [46]. |
| Anti-B Domain Antibodies | Polyclonal or monoclonal antibodies targeting the G5-E repeats of Aap/SasG. | Blocks intercellular interactions and inhibits biofilm formation [46]. |
| Recombinant B-Domain Protein | Purified protein corresponding to the G5-E repeat region of Aap or SasG. | Used for in vitro dimerization studies and as a competitive inhibitor in biofilm assays [46]. |
The discovery and characterization of proteinaceous biofilm accumulation mechanisms represent a critical evolution in our understanding of staphylococcal pathogenesis. The Zn²⁺-dependent "zipper" model of Aap and SasG provides a elegant mechanistic explanation for PIA-independent intercellular adhesion. This knowledge reframes the biofilm paradigm from one centered on a single polysaccharide to a multifaceted model where diverse molecular strategies, including protein-based matrices, can lead to the same pathogenic outcome. This complexity is further underscored by genomic evidence suggesting that different Staphylococcus species have independently evolved different mechanisms for biofilm production multiple times throughout evolution [44].
For researchers and drug development professionals, this expanded understanding opens new avenues for therapeutic intervention. Targeting the proteolytic activation of Aap, disrupting the Zn²⁺-binding site with specific chelators or small molecules, or using recombinant B domains as decoy receptors are all potential strategies under investigation. Combating staphylococcal biofilms will likely require a multi-pronged approach that neutralizes both polysaccharide and proteinaceous matrix components, and a deep understanding of the mechanisms detailed in this whitepaper is the essential first step toward that goal.
Within the context of polysaccharide intercellular adhesin (PIA) research in Staphylococcus, the icaADBC locus is recognized as the genetic cornerstone for biofilm formation, a key virulence mechanism in device-related infections [1] [2]. The operon encodes enzymes for the synthesis of PIA, a poly-β(1-6)-N-acetylglucosamine (PNAG) that facilitates cell-cell adhesion and forms a critical structural component of the staphylococcal biofilm matrix [11]. While detecting the ica locus through molecular methods is straightforward, correlating its mere presence with the clinical infectivity of a strain presents a significant diagnostic challenge. This whitepaper delves into the complexities underlying this discrepancy, arguing that a sophisticated understanding of ica regulation and expression is paramount for developing accurate diagnostic tools and effective anti-biofilm therapeutics for research and drug development.
The Core Diagnostic Challenge: Presence Versus Expression
The fundamental challenge in clinical diagnostics is that the detection of the ica locus does not reliably predict biofilm-forming phenotype or clinical infectivity. Multiple studies have demonstrated a disconnect between genetic potential and observable behavior in vitro.
Clinical Evidence of the Disconnect
Table 1: Representative Clinical Studies Highlighting the ica-Phenotype Discordance
| Study Context | Finding on ica Locus Presence | Finding on Biofilm Formation/PIA Production | Implication |
|---|---|---|---|
| S. aureus from prosthetic joint infections [47] [48] | icaA gene was present in all 15 clinical isolates. | Only one isolate weakly produced biofilm in vitro; none showed hemagglutination (a PIA-associated trait). | PIA expression may be tightly restricted to specific in vivo conditions. |
| Clinical isolates of S. epidermidis [49] | The entire icaADBC operon was present in 45% of isolates from catheter-related and other nosocomial infections. | The presence of the entire genetic cluster did not always correlate with biofilm production under different culture conditions. | Diagnostic assays must account for regulatory mechanisms beyond gene presence. |
| Device-related animal model study [10] | S. aureus strains (e.g., Newman) contained the ica locus but were PIA-negative after standard in vitro growth. | PIA and ica-specific transcripts became detectable in vivo, culminating in biofilm formation by day 8 of infection. | The in vivo environment provides strong inducing signals absent in standard labs. |
Molecular Mechanisms Underpinning the Challenge
The discrepancy between gene presence and phenotype is not due to chance but is a consequence of multifaceted molecular regulation.
- Tight Genetic Regulation: The expression of the icaADBC operon is controlled by a complex network of regulatory factors. These include the dedicated repressor IcaR, as well as global regulators like SarA and the alternative sigma factor SigB (σB) [50]. This network integrates a variety of environmental signals, making ica expression highly context-dependent.
- Phase Variation: Both S. aureus and S. epidermidis can undergo phase variation for biofilm production. This reversible, high-frequency switching can be caused by the insertion of an insertion sequence (IS) element within the icaC gene or by the expansion and contraction of a tandem repeat sequence naturally present in icaC [1] [2]. This mechanism can spontaneously generate biofilm-negative subpopulations from a biofilm-positive, ica-positive parent strain.
- ica-Independent Biofilm Pathways: Some staphylococcal strains form biofilms primarily via proteinaceous components rather than PIA. These mechanisms can involve surface proteins like Bap (biofilm-associated protein) in S. aureus or Aap (accumulation-associated protein) in S. epidermidis [50] [11]. In these strains, the presence or absence of the ica locus is not the primary determinant of biofilm-mediated infectivity.
Diagram 1: Relationship between ica locus presence and clinical infectivity. The pathway shows that ica presence is merely the starting point in a complex network of regulatory and alternative mechanisms that ultimately determine biofilm formation and infectivity.
Essential Methodologies for Advanced Correlation Studies
To move beyond simple genetic detection, researchers must employ a multi-faceted experimental approach that captures both the genetic potential and the functional expression of biofilm-related virulence.
Genotypic Detection Protocols
The foundational step is confirming the presence of the ica operon. The standard protocol involves PCR amplification of key genes.
- Target Genes: A comprehensive analysis should target multiple genes within the operon, notably icaA and icaD, which work in concert as the N-acetylglucosaminyltransferase, and are essential for PIA synthesis [20] [11].
- Primer Design: Primers should be designed to amplify internal fragments of these genes (e.g., a 419-bp product for icaA [47]).
- Procedure: Standard PCR is performed on extracted bacterial genomic DNA. The amplicons are then separated by gel electrophoresis and visualized to confirm the presence and size of the expected products.
Phenotypic Biofilm Assays
Genotypic data must be correlated with robust phenotypic assays for biofilm formation.
Microtiter Plate (MtP) Assay [20] [2]:
- Culture Preparation: Grow test strains overnight in Tryptic Soy Broth (TSB) supplemented with a inducing agent such as 0.25% glucose.
- Inoculation and Incubation: Dilute cultures 1:200 in fresh medium and incubate statically in U-bottom polystyrene microtiter plates at 37°C for 24 hours.
- Washing and Staining: Gently wash the wells twice with phosphate-buffered saline (PBS) to remove non-adherent cells. Adherent biofilms are then fixed and stained with 0.1% safranin or crystal violet.
- Quantification: For crystal violet, elute the bound dye with ethanol or acetic acid and measure the optical density at a specific wavelength (e.g., 570 nm) using a microtiter plate reader. This provides a quantitative measure of biofilm biomass.
PIA Detection via Immunoassay [20] [10]:
- Antigen Preparation: Grow bacteria in inducing medium (e.g., TSB + 1% glucose, potentially under anaerobic conditions). Cells are harvested, resuspended in 0.5 M EDTA, and boiled to extract surface polysaccharides.
- Proteinase K Treatment: The supernatant is treated with proteinase K to degrade any contaminating proteins that could cause non-specific antibody binding.
- Immunodetection: The extract is spotted onto a nitrocellulose membrane and probed with a specific anti-PIA antibody (e.g., absorbed polyclonal rabbit IgG). Bound antibody is detected using a secondary enzyme-conjugated antibody and a colorimetric or chemiluminescent substrate.
Transcriptional Analysis
Linking genotype to phenotype requires assessing the expression of the ica operon.
- RNA Extraction: Extract total RNA from bacteria grown under conditions mimicking the in vivo environment (e.g., from animal model exudates or after in vitro induction with high salt, ethanol, or anaerobic growth) [10].
- Quantitative Reverse Transcription PCR (qRT-PCR): This is the gold standard for quantifying gene expression. Using primers specific for icaA or icaD, and a housekeeping gene for normalization, researchers can precisely measure ica operon transcription levels. This technique is sensitive enough to detect upregulation in response to specific cues, such as the presence of other bacterial species in a polymicrobial biofilm [51].
Diagram 2: Experimental workflow for comprehensive analysis. A multi-pronged approach is necessary to establish a reliable correlation between the ica locus and infectivity, integrating genotypic, phenotypic, and transcriptional data.
The Scientist's Toolkit: Key Research Reagent Solutions
Table 2: Essential Reagents and Assays for Staphylococcal Biofilm Research
| Reagent/Assay | Function/Application | Key Consideration |
|---|---|---|
| Anti-PIA/PNAG Antibody | Specific detection of PIA in immunoassays (e.g., dot blot, immunofluorescence) and validation of deacetylated PNAG [20] [10]. | Antibody should be pre-absorbed to ensure specificity. Critical for distinguishing PIA-dependent biofilms. |
| icaADBC-specific PCR Primers | Genotypic screening for the presence of the operon and its individual genes in clinical isolates [20] [47]. | Targeting multiple genes (e.g., icaA and icaD) increases detection reliability. |
| qRT-PCR Assay Kits | Quantitative analysis of ica operon transcription under different environmental or co-culture conditions [10] [51]. | Essential for studying regulation; requires careful RNA preservation and normalization. |
| Inducing Media (TSB + Glucose/NaCl) | In vitro induction of ica expression and PIA production for phenotypic assays [20] [50]. | Standard TSB may not induce biofilm; supplements like 1% glucose are often necessary. |
| Dispersin B | Specific glycoside hydrolase that degrades PNAG. Used as a tool to confirm the structural role of PIA in a biofilm [2]. | A key experimental control to validate PIA-dependent biofilm phenotypes. |
| Microtiter Plates (Polystyrene) | Standardized platform for high-throughput quantification of biofilm formation [20] [2]. | The material and surface treatment (e.g., U-bottom) can influence attachment. |
The diagnostic challenge of correlating the ica locus with clinical infectivity stems from the complex biology of staphylococcal biofilms. The presence of the ica genes is a necessary but insufficient predictor of virulence. True predictive power lies in understanding and measuring the expression of this locus, which is governed by a dense network of regulatory inputs and subject to phase variation. For researchers and drug developers, this underscores the limitation of diagnostics based solely on genetic presence. The future lies in integrating advanced techniques—such as transcriptional profiling directly from clinical specimens and developing sensitive in vivo imaging for PIA—that capture the dynamic expression of virulence. Furthermore, the existence of ica-independent biofilm mechanisms necessitates a poly-therapeutic approach to combat biofilm-related infections, targeting not only PIA but also protein and eDNA components of the matrix. Overcoming these diagnostic challenges is crucial for accurately identifying high-risk bacterial strains, evaluating novel anti-biofilm compounds, and ultimately improving patient outcomes in device-related infections.
The pathogenesis of Staphylococcus aureus and Staphylococcus epidermidis is critically dependent on their ability to form biofilms, particularly in the context of implant-associated infections. The extracellular matrix of staphylococcal biofilms is a key virulence component, providing structural integrity and conferring resistance to host immune defenses and antibiotics. A principal constituent of this matrix is polysaccharide intercellular adhesin (PIA), a cationic, partially deacetylated polymer of β-1,6-linked N-acetylglucosamine (poly-β(1-6)-N-acetylglucosamine; PNAG) [3] [2]. The synthesis of PIA is mediated by the icaADBC operon, and its production is strongly influenced by environmental conditions [52] [3]. Beyond its role as an adhesin, PIA is a crucial shield, protecting staphylococci from major components of the human innate immune system, including antimicrobial peptides and phagocytosis by polymorphonuclear leukocytes [53].
Staphylococcal biofilm development is not a constitutive process but is intricately regulated. The accessory gene regulator (agr) quorum sensing (QS) system serves as a central coordinator of virulence, enabling bacterial populations to communicate and synchronize their behavior based on cell density [54] [55]. The agr system typically upregulates the production of secreted toxins and exoenzymes while repressing the expression of surface proteins [54]. Intriguingly, the relationship between the agr system and biofilm formation is complex and appears to be antagonistic; QS activity can promote the dispersal of bacteria from a mature biofilm, facilitating the spread to new infection sites [2]. This positions the agr system as a key target for quorum quenching (QQ), a therapeutic strategy that interferes with bacterial cell-to-cell communication to combat biofilm-related infections [2] [56]. This technical guide explores the molecular basis of QS interference in the context of PIA-dependent staphylococcal biofilms, providing detailed methodologies and data analysis for researchers in the field.
Molecular Mechanisms: The agr System and PIA Biosynthesis
The agr Quorum Sensing System
The agr locus is composed of two primary transcriptional units, RNAII and RNAIII, driven by the P2 and P3 promoters, respectively [54] [55]. The RNAII operon encodes the core components of the QS signaling circuit:
- AgrD: The precursor of the autoinducing peptide (AIP).
- AgrB: A transmembrane protein that processes and exports AgrD, modifying it into a cyclic thiolactone AIP.
- AgrC: A histidine kinase sensor that binds the extracellular AIP.
- AgrA: A response regulator that, when activated by phosphoryl transfer from AgrC, enhances transcription from the P2 and P3 promoters [54] [55].
At high cell densities, the accumulated AIP activates the AgrCA two-component system, leading to a rapid increase in the transcription of RNAIII. RNAIII is the actual effector of the agr response, acting as a regulatory RNA that modulates the translation and transcription of a vast regulon of virulence genes [54]. This system allows staphylococci to shift from a colonizing phenotype (expressing surface adhesins) to an invasive, dispersive phenotype (expressing secreted toxins) [54] [55].
Biosynthesis and Regulation of Polysaccharide Intercellular Adhesin (PIA)
PIA biosynthesis is catalyzed by enzymes encoded by the icaADBC operon [3] [2]:
- IcaA: An N-acetylglucosaminyltransferase that synthesizes PIA oligomers from UDP-N-acetylglucosamine. It has low activity on its own.
- IcaD: Works synergistically with IcaA to form an efficient synthetase complex, enhancing the transferase activity.
- IcaC: Believed to be involved in the externalization and elongation of the nascent polysaccharide chain.
- IcaB: An N-deacetylase that partially deacetylates PIA, imparting a positive charge essential for its adhesive properties and role in immune evasion [3] [2].
The expression of the ica operon is controlled by a network of regulatory inputs. The operon is repressed by IcaR, a DNA-binding protein that binds upstream of the icaA transcriptional start site [52] [3]. Furthermore, environmental signals such as nutrient availability, low oxygen, high osmolarity, and elevated temperatures can induce PIA production, often through regulators like the alternative sigma factor σB [52] [2]. A crucial connection to central metabolism exists: conditions that repress the tricarboxylic acid (TCA) cycle (e.g., low iron, anaerobic growth) strongly promote PIA production [52].
The Critical Interaction: agr-Mediated Repression of Biofilm
The agr system and PIA-mediated biofilm formation exist in a state of reciprocal inhibition, a key conceptual point for QQ strategies. While environmental signals can favor PIA production and biofilm establishment, the activation of the agr system at high cell density can trigger biofilm dispersal [2]. This dispersal is thought to be part of a strategic cycle where bacteria within a biofilm eventually escape to disseminate the infection. Therefore, targeting the agr system via QQ does not aim to bolster biofilms but rather to lock the bacterial population in a non-dispersing, potentially more treatable state by preventing the QS-controlled switch to a dispersive phenotype.
Diagram 1: Regulatory network of PIA biosynthesis and agr quorum sensing in staphylococci. The diagram illustrates the opposing relationship between the PIA-mediated biofilm formation pathway (green) and the agr-mediated quorum sensing pathway (blue). Environmental signals (yellow) influence both pathways. The agr system, via its effector RNAIII, inhibits the icaADBC operon and promotes biofilm dispersal, creating a key target for quorum quenching interventions.
Experimental Analysis of Quorum Quenching Efficacy
Key Quantitative Findings on agr and Biofilm Phenotype
The complex relationship between the agr system, PIA production, and biofilm formation has been quantified in several key studies. The data below summarize the phenotypic changes observed when the agr system is disrupted, either genetically or via quorum quenching agents.
Table 1: Impact of agr Disruption on Staphylococcal Biofilm and Virulence Factors
| Parameter Measured | Experimental Model | Effect of agr Deletion or Inhibition | Citation |
|---|---|---|---|
| Biofilm Formation | S. epidermidis agr deletion mutant | Increased biofilm formation and primary attachment | [57] |
| PIA Expression | S. epidermidis agr deletion mutant | No significant change in PIA levels | [57] |
| Autolysin (AtlE) Expression | S. epidermidis agr deletion mutant | Increased expression | [57] |
| δ-toxin Production | S. epidermidis agr deletion mutant | Absent | [57] |
| Primary Attachment | S. epidermidis with cross-inhibiting AIPs | Mimicked agr mutation, significantly enhanced biofilm | [57] |
| Phagocytosis by PMNs | S. epidermidis PIA-negative mutant | Significantly increased compared to wild-type | [53] |
| Susceptibility to Antimicrobial Peptides | S. epidermidis PIA-negative mutant | Significantly increased susceptibility to β-defensin 3, LL-37, dermcidin | [53] |
The data in Table 1 reveals a critical nuance: the hyper-biofilm phenotype of agr mutants is not necessarily due to increased PIA production but involves other factors like enhanced primary attachment mediated by autolysin AtlE [57]. This underscores that QQ's anti-dispersal effect is multifaceted. Furthermore, the susceptibility data highlights a key therapeutic synergy: disrupting QS to maintain a biofilm state could render bacteria more vulnerable to clearance by the innate immune system, especially when PIA is targeted simultaneously [53].
Detailed Experimental Protocol: Assessing QQ Agents on Biofilm Formation
The following is a standardized protocol for evaluating the efficacy of potential quorum-quenching compounds against staphylococcal biofilms, incorporating key methodologies from the search results.
Method: In Vitro Biofilm Assay with Quorum Quenching Compounds
1. Bacterial Strains and Growth Conditions
- Strains: Use relevant biofilm-forming strains (e.g., S. epidermidis O-47, S. aureus SA113) and their isogenic mutants (e.g., S. epidermidis Δagr, S. aureus SA113Δica) as controls [57] [58].
- Culture Media: Tryptic Soy Broth (TSB) supplemented with 0.5% - 1% glucose (TSBg) to enhance PIA production [52] [58].
- Growth Conditions: Inoculate bacteria from overnight cultures and grow to mid-exponential phase (OD₆₀₀ ~ 0.5) under aerobic conditions with shaking at 160 rpm [52].
2. Treatment with Quorum Quenching Compounds
- QQ Compounds: Prepare stock solutions of candidate inhibitors (e.g., synthetic AIP analogs, cross-inhibiting AIPs from other agr groups [57], or small-molecule inhibitors).
- Experimental Setup: In a sterile 96-well polystyrene microtiter plate, add 180 µL of diluted bacterial suspension (e.g., 1:200 in TSBg) to each well.
- Compound Addition: Add 20 µL of the QQ compound at varying concentrations (e.g., 1-100 µM). Include controls:
- Negative Control: Bacteria with vehicle (e.g., DMSO, water).
- Positive Control: Bacteria with a known anti-biofilm agent (e.g., Dispersin B, DNase I [2]).
- Sterility Control: TSBg only.
- Incubation: Incubate the plate statically for 24 hours at 37°C to allow for biofilm development.
3. Biofilm Quantification (Safranin Staining)
- Carefully remove the planktonic cells and growth medium from each well.
- Wash the adhered biofilms gently twice with 200 µL of phosphate-buffered saline (PBS) to remove non-adherent cells.
- Fix the biofilms by air-drying the plate for approximately 1 hour.
- Stain the biofilms with 200 µL of a 0.1% (w/v) safranin solution for 15-30 minutes.
- Remove the stain and wash the wells thoroughly with water to remove excess dye.
- Elute the bound safranin from the stained biofilms by adding 200 µL of 30% (v/v) acetic acid and incubating for 15 minutes with shaking.
- Transfer 100 µL of the eluted dye to a new microtiter plate and measure the absorbance at 492 nm using a plate reader [58].
4. Complementary Analyses
- PIA Quantification: Extract PIA from biofilms grown in parallel using 0.5 M EDTA (pH 8.0) at 100°C. Quantify PIA via immunoblotting with specific anti-PIA antiserum [52].
- Viability Counts: Perform colony-forming unit (CFU) counts on treated and untreated biofilms after sonication to determine if QQ effects are bactericidal or specifically anti-biofilm.
- Gene Expression: Use RT-qPCR to analyze the transcription of key genes (e.g., icaA, icaR, hld [within RNAIII], agrA) in treated vs. untreated biofilms [57].
5. Data Interpretation
- A successful QQ compound will typically show a dose-dependent reduction in biofilm biomass (safranin staining) without a significant reduction in planktonic viability, indicating an anti-biofilm而非 bactericidal mechanism.
- Effective QQ should lead to downregulation of RNAIII and potentially upregulation of icaADBC, confirming the interference with the agr system and a shift towards the biofilm phenotype.
- The use of isogenic mutants (e.g., Δagr, Δica) is crucial to confirm that the compound's mechanism of action is specifically tied to the agr pathway and PIA production.
The Scientist's Toolkit: Key Research Reagents
Table 2: Essential Reagents for Studying Quorum Quenching and PIA-Mediated Biofilms
| Reagent / Tool | Function / Description | Key Application in Research |
|---|---|---|
| Isogenic Mutant Strains (e.g., Δagr, Δica) | Strains with specific gene deletions in QS or PIA biosynthesis pathways. | Serves as essential controls to validate the specificity of QQ compounds and delineate molecular pathways [57] [58]. |
| Synthetic Autoinducing Peptides (AIPs) | Chemically synthesized peptides matching native AIPs or inhibitory analogs. | Used to exogenously manipulate the agr system; cross-inhibiting AIPs can mimic QQ [57]. |
| Anti-PIA Antiserum | Polyclonal antibodies raised against purified PIA/PNAG. | Essential for quantifying PIA production via immunoblotting and visualizing its localization in the biofilm matrix [52] [53]. |
| Dispersin B | A glycoside hydrolase enzyme that cleaves β-1,6-linked PIA/PNAG. | Used as a positive control for biofilm disruption and to study the structural role of PIA [2]. |
| Fluorocitrate | A specific inhibitor of the TCA cycle enzyme aconitase. | Tool to investigate the link between metabolic status (TCA cycle repression) and increased PIA production [52]. |
| Microtiter Plate (MtP) Biofilm Assay | A high-throughput, static method for growing and quantifying biofilms. | The standard first-line assay for screening anti-biofilm and QQ efficacy [2] [58]. |
| Proteinase K & DNase I | Enzymes that degrade protein and extracellular DNA (eDNA) in the biofilm matrix. | Used to dissect the composition of the biofilm and study synergistic effects with QQ agents [2]. |
The strategic interference with staphylococcal quorum sensing represents a paradigm shift in anti-biofilm therapy. By targeting the agr system through quorum quenching, it is possible to disrupt the bacterial communication that orchestrates virulence and biofilm dispersal. The intricate, often antagonistic, relationship between the agr system and PIA-dependent biofilm formation provides a compelling therapeutic window. The experimental frameworks and tools detailed in this guide provide a foundation for the discovery and validation of novel QQ agents. Future work in this field will likely focus on combining QQ strategies that prevent dispersal with direct matrix-degrading agents like Dispersin B, which targets PIA, and conventional antibiotics. This multi-pronged approach, informed by a deep understanding of the underlying molecular biology, holds significant promise for overcoming the formidable challenge of chronic, biofilm-associated staphylococcal infections.
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), plays a complex and variable role in staphylococcal prosthetic joint infections (PJIs). While long recognized as a key component of the staphylococcal biofilm matrix, contemporary research reveals that its expression and functional significance are highly context-dependent, influenced by genetic regulation, environmental conditions, and bacterial lineage. This technical review synthesizes current molecular and transcriptomic evidence to delineate the factors governing PIA production in vivo, its correlation with clinical outcomes, and the resultant implications for diagnostic and therapeutic strategies. The emerging paradigm shift acknowledges that PIA-independent biofilm mechanisms are equally critical in many clinical isolates, necessitating a refined approach to combat these resilient infections.
Prosthetic joint infection (PJI) remains one of the most devastating complications in orthopedics, with staphylococcal species, predominantly Staphylococcus aureus and Staphylococcus epidermidis, being the most frequently isolated pathogens [59] [60]. The therapeutic recalcitrance of these infections is largely attributed to the formation of bacterial biofilms—structured communities of bacteria encased in a self-produced extracellular matrix on the implant surface [61] [60].
The polysaccharide intercellular adhesin (PIA), a poly-β(1-6)-N-acetylglucosamine (PNAG), has been extensively studied as a primary molecular component responsible for intercellular adhesion and biofilm accumulation in staphylococci [1] [2]. Its synthesis is mediated by the icaADBC operon, which is functionally conserved across many staphylococcal species [62]. However, within the specific microenvironment of a prosthetic joint, the expression and functional contribution of PIA are not absolute. This whitepaper examines the differential expression of PIA in PJI sites, framing it within the broader thesis that a comprehensive understanding of this variability is paramount for advancing diagnostic and anti-biofilm therapeutic interventions.
Molecular Genetics and Regulation of PIA Synthesis
TheicaADBCOperon and PIA Biosynthesis
The synthesis of PIA is a multi-step process directed by the icaADBC operon. Each gene product plays a distinct role in the synthesis, export, and modification of the final polysaccharide structure.
Table 1: Gene Components of the icaADBC Operon and Their Functions
| Gene | Function of Gene Product |
|---|---|
| icaA | N-acetylglucosaminyltransferase; synthesizes PIA oligomers from UDP-N-acetylglucosamine [1] [2]. |
| icaD | Stabilizes and enhances the activity of IcaA, providing optimal efficiency for PIA synthesis [1] [2]. |
| icaC | Involved in the externalization and full elongation of the nascent polysaccharide chain [1] [2]. |
| icaB | N-deacetylase; responsible for the partial deacetylation of PIA, which confers a positive charge and is critical for its adhesive properties [1] [2]. |
The product of this operon is a partially deacetylated, positively charged polysaccharide that facilitates bacterial aggregation and strong attachment to surfaces, forming the structural scaffold of the biofilm matrix [2] [62].
Regulatory Networks InfluencingicaADBCExpression
The expression of the ica locus is not constitutive but is instead influenced by a complex network of regulatory systems that respond to environmental conditions, such as those found in the PJI niche.
Diagram 1: Regulatory network of the icaADBC operon. The sarA protein acts as a positive regulator, while IcaR and the agr system typically repress PIA production. All are influenced by local environmental conditions.
The staphylococcal accessory regulator A (sarA) is a key positive regulator of biofilm formation. It upregulates the transcription of the icaADBC operon and also represses proteases that could degrade proteinaceous biofilm components, thereby functioning through both PIA-dependent and PIA-independent pathways [61]. In contrast, the accessory gene regulator (agr) quorum sensing system often functions as a negative regulator of biofilm formation. It promotes the dispersal phase of the biofilm lifecycle, favoring the spread of bacteria to new infection sites. Consequently, spontaneous agr mutants are frequently selected in vivo, leading to a more stable biofilm phenotype [61] [2]. Another critical regulator is IcaR, encoded by a gene located upstream of icaA, which directly represses the icaADBC operon [62]. The expression of these regulators is, in turn, sensitive to local environmental conditions, such as oxygen tension, osmotic stress, and the presence of antibiotics [1] [2].
Evidence of PIA's Variable Expression in Prosthetic Joint Infections
Genetic and Phenotypic Prevalence in Clinical Isolates
The presence of the ica locus does not always correlate with biofilm-forming capacity in vivo, underscoring the complexity of its regulatory control. A 2025 study on synovial fluid isolates from PJI patients found a statistically significant difference in c-di-GMP production (a key secondary messenger that promotes biofilm formation) between strains harboring the icaA gene and those without it (p = 0.016) [59]. This suggests a functional link between the genetic potential for PIA production and the broader biofilm regulatory circuitry. The same study identified a diverse array of biofilm-associated genes beyond the ica operon, including clfA and fnbA in S. aureus, and atlE and fbe in coagulase-negative staphylococci (CoNS) [59].
Table 2: Biofilm-Associated Genes in Staphylococcal PJI Isolates (2025 Study)
| Species | Biofilm-Associated Genes Identified | Significant Correlation |
|---|---|---|
| S. aureus | icaA, clfA, fnbA |
Significant linear regression between biofilm formation and c-di-GMP production (p=0.016) [59]. |
| Coagulase-Negative Staphylococci (CoNS) | icaA, atlE, fbe |
Oxacillin resistance detected more frequently in strains carrying the fbe gene (p=0.031) [59]. |
Transcriptomic Profiles:In Vivovs.In VitroConditions
Advanced transcriptomic analyses have provided unprecedented insights into the adaptive responses of staphylococci within the PJI environment. RNA sequencing (RNA-seq) of S. aureus from sonicate fluid of explanted prosthetic devices reveals how gene expression shifts dramatically from laboratory conditions to the human host.
A pivotal finding is the upregulation of ica-independent biofilm formation genes in vivo. These include fibronectin-binding proteins (fnb), serine-aspartate repeat proteins (sdr), and the elastin binding protein (ebpS) [63]. This indicates a switch towards a protein-based biofilm strategy in the actual infection niche, which may compensate for or replace the PIA-mediated mechanism. Furthermore, transcriptomic profiling of S. epidermidis from PJIs shows significant upregulation of metal sequestration systems in vivo, such as the staphylopine activity genes (cntA, cntK, cntL, cntM), highlighting a key metabolic adaptation that occurs alongside biofilm formation [64]. These in vivo transcriptomic signatures confirm that the biofilm phenotype in PJIs is a composite of multiple mechanisms, with PIA's role being highly variable and strain-specific.
Experimental Models and Methodologies for Investigating PIA
Protocol: Evaluating Biofilm Formation and PIA Production
Method: Microtiter Plate (MtP) Biofilm Assay with Crystal Violet Staining [59] [2].
- Procedure:
- Grow bacterial isolates in a suitable broth (e.g., Tryptic Soy Broth with 1% glucose) in 96-well polystyrene plates.
- Incubate for 24-72 hours at 37°C under static conditions.
- Carefully remove planktonic cells and wash the adherent biofilms with phosphate-buffered saline (PBS).
- Fix the biofilm with 99% methanol or ethanol for 15 minutes.
- Stain with 0.1% crystal violet solution for 5-20 minutes.
- Wash extensively to remove unbound dye.
- Elute the bound crystal violet with 33% glacial acetic acid.
- Measure the optical density (OD) of the eluent at 570-595 nm to quantify the total adhered biofilm biomass.
- Application: This high-throughput method allows for the phenotypic screening of biofilm-forming capacity across a large number of clinical isolates. It is often coupled with genetic analysis (PCR for ica genes) to correlate genotype with phenotype [2].
Protocol: RNA Sequencing forIn VivoGene Expression Analysis
Method: Total RNA-Sequencing (RNA-seq) from Clinical Specimens [63] [65].
- Procedure:
- Sample Collection: Obtain sonicate fluid from explanted prosthetic hardware or synovial fluid during revision surgery.
- RNA Stabilization & Isolation: Immediately stabilize RNA (e.g., with RNAlater) or snap-freeze samples at -80°C. Total RNA is isolated using kits designed for complex samples (e.g., miRNeasy Serum/Plasma Kit), followed by removal of host and bacterial ribosomal RNA.
- Library Preparation & Sequencing: Construct cDNA libraries using a system that accommodates low-input RNA (e.g., Ovation SoLo RNA-Seq System). Sequence on a high-throughput platform (e.g., Illumina HiSeq).
- Bioinformatic Analysis:
- Remove host-derived reads using BioBloom Tools [63].
- Map remaining reads to a reference or pan-genome of the bacterial isolate using tools like Kallisto [63].
- Perform differential expression analysis (e.g., using Sleuth) to compare transcript levels between in vivo samples and matched in vitro cultures [63].
- Application: This protocol directly captures the transcriptional state of bacteria during human infection, revealing the in vivo expression of the ica locus and its regulators, as well as alternative biofilm pathways [63] [65] [64].
Protocol: Mimicking the PJI EnvironmentIn Vitro
Method: Modified Bone-Like Environment (BLE+) Model on Suspended Titanium Pegs [66].
- Procedure:
- Culture Conditions: Grow clinical S. aureus strains in BLE+ medium under hypoxic conditions (2.5% O₂) to simulate the bone site environment.
- Biofilm Support: Use suspended titanium pegs to hold bacteria in the medium, specifically driving active bacterial adhesion and minimizing interference from sedimented aggregates.
- Incubation & Renewal: Incubate for 72 hours. Renew the culture medium after 8 hours of initial adhesion to limit the interaction between planktonic bacteria and the developing biofilm.
- Analysis: Assess biofilm formation by quantifying both planktonic and adherent bacteria (via colony-forming units, CFU) and total biomass (via crystal violet staining).
- Application: This model provides a more clinically relevant platform for studying biofilm formation and testing anti-biofilm agents by incorporating critical parameters like hypoxia, a relevant growth medium, and a prosthetic surface [66].
Diagram 2: Integrated workflow for investigating PIA's role in PJI, combining genotypic, phenotypic, and transcriptomic approaches.
The Scientist's Toolkit: Key Research Reagents and Materials
Table 3: Essential Reagents for Investigating PIA and Biofilm in PJI
| Reagent / Material | Function in Research | Example Use Case |
|---|---|---|
| Crystal Violet | A basic dye that binds nonspecifically to negatively charged surface molecules and polysaccharides in the biofilm matrix. | Quantification of total biofilm biomass in microtiter plate assays [59] [66]. |
| Titanium Pegs / Plates | Provides a physiologically relevant abiotic surface (mimicking orthopedic implants) for studying bacterial adhesion and biofilm formation. | In vitro biofilm models using orthopedic implant materials [66]. |
| Sonicate Fluid (from Explanted Prostheses) | The sample matrix containing bacteria dislodged from the biofilm on the implant surface; considered the gold standard for PJI diagnosis and research. | Source for transcriptomic analysis (RNA-seq) of in vivo bacterial gene expression [63] [65]. |
| RNAlater / Snap-Freezing | Chemical or physical methods to immediately stabilize RNA and preserve the in vivo transcriptome at the time of sample collection. | Preserving bacterial RNA from clinical samples for subsequent RNA sequencing [63]. |
| Ribo-Zero rRNA Removal Kit (Bacteria) | Removes the abundant ribosomal RNA (rRNA) from total RNA samples, enriching for messenger RNA (mRNA) to improve sequencing depth. | Preparation of RNA-seq libraries from bacterial samples [63]. |
| BLE+ Medium | A customized culture medium designed to more closely mimic the nutritional and physicochemical environment of a bone and joint infection site. | Culturing bacteria under conditions that promote in vivo-like biofilm phenotypes [66]. |
The role of PIA in prosthetic joint infections is defined by its variability. Its expression is not a binary determinant of virulence but is one component in a sophisticated, adaptable biofilm program regulated by a network of genetic and environmental factors. The evidence is clear: while the icaADBC operon is a significant player, successful bacterial colonization and persistence in the PJI niche often involve redundant, ica-independent mechanisms, such as proteinaceous adhesins and regulated metabolic shifts.
This understanding mandates a shift in therapeutic development. Targeting a single molecule like PIA is unlikely to yield broad-spectrum efficacy. Future strategies must consider the following approaches:
- Targeting Master Regulators: Focusing on central regulatory systems like sarA, which controls both PIA-dependent and independent pathways, could disrupt biofilm integrity more effectively [61].
- Anti-Virulence Therapies: Developing quorum-quenching compounds or inhibitors of key alternative adhesins (e.g., FnBPs) could prevent biofilm formation without exerting direct lethal pressure that drives resistance [2].
- Vaccine Development: The partially deacetylated form of PNAG is a promising vaccine candidate, as it is retained on the bacterial surface and can promote opsonophagocytosis, offering a potential prophylactic solution [1] [2].
Ultimately, overcoming the challenge of staphylococcal PJIs will require a multi-pronged strategy informed by a deep understanding of the differential expression and functional redundancy of biofilm components like PIA, both in the laboratory and within the human host.
Biofilm-associated infections represent a significant challenge in clinical settings, primarily due to their enhanced tolerance to antimicrobial agents. A key component of the staphylococcal biofilm matrix is polysaccharide intercellular adhesin (PIA), also known as poly-β(1-6)-N-acetylglucosamine (PNAG). This review provides an in-depth technical analysis of the disconnect between standard minimum inhibitory concentration (MIC) assays and the more clinically relevant minimum biofilm eradication concentration (MBEC) evaluations. We explore the structural and functional roles of PIA in conferring this tolerance, detail experimental methodologies for quantifying biofilm-specific antibiotic efficacy, and discuss emerging therapeutic strategies that target PIA. The data synthesized herein underscore the critical need for biofilm-specific testing protocols in both diagnostic and drug development workflows to effectively address persistent staphylococcal infections.
The Clinical Problem of Biofilm-Associated Infections
Bacterial biofilms are structured communities of cells encased in an extracellular matrix and attached to a surface or interface. In clinical contexts, biofilms are implicated in approximately 65-80% of bacterial infections, particularly those associated with medical implants such as catheters and prosthetic joints [18]. Bacteria within biofilms can resist antibiotics at concentrations up to 1000 times higher than those effective against their planktonic (free-floating) counterparts, leading to chronic, recalcitrant infections [2]. This remarkable tolerance is not primarily due to genetic resistance mechanisms but rather to the physical and physiological structure of the biofilm itself.
Polysaccharide Intercellular Adhesin: A Key Biofilm Matrix Component
In staphylococci, particularly Staphylococcus epidermidis and Staphylococcus aureus, the exopolysaccharide polysaccharide intercellular adhesin (PIA) is a crucial structural component of the biofilm matrix [3] [1]. PIA is a linear polymer of β-1,6-linked N-acetylglucosamine that is partially deacetylated (approximately 15-20%), giving it a positive charge [3] [2]. This chemical composition is critical to its function. The biosynthesis of PIA is mediated by the icaADBC operon, with IcaA serving as an N-acetylglucosaminyltransferase, IcaD enhancing IcaA's efficiency, IcaC involved in externalization, and IcaB functioning as an N-deacetylase responsible for the partial deacetylation essential for PIA's adhesive properties [1] [2].
Table 1: Genetic Components of PIA Biosynthesis in Staphylococci
| Gene | Function | Role in Biofilm Formation |
|---|---|---|
| icaA | N-acetylglucosaminyltransferase | Synthesizes PIA oligomers from UDP-N-acetylglucosamine |
| icaD | Regulatory subunit | Optimizes efficiency of IcaA |
| icaC | Membrane transporter | Involved in externalization of nascent polysaccharide |
| icaB | N-deacetylase | Partially deacetylates PIA, essential for adhesion and biofilm integrity |
MIC vs. MBEC: Fundamental Concepts and Methodological Approaches
Minimum Inhibitory Concentration: Planktonic Cell Focus
The Minimum Inhibitory Concentration (MIC) is defined as the lowest concentration of an antimicrobial agent that prevents visible growth of a planktonic bacterial isolate under standardized conditions. MIC testing represents the current gold standard in clinical antimicrobial susceptibility testing but fails to account for the altered physiology and protective environment of biofilm-embedded cells.
Minimum Biofilm Eradication Concentration: Biofilm Relevance
The Minimum Biofilm Eradication Concentration (MBEC) is defined as the lowest concentration of an antimicrobial agent that eradicates a mature biofilm, typically measured using assays that quantify metabolic activity or viable counts following antibiotic exposure [67] [2]. The MBEC assay system typically involves growing biofilms on pegs or surfaces, exposing them to antimicrobial agents, and then determining the concentration required to kill the biofilm-embedded cells.
Quantitative Disparities Between MIC and MBEC
Substantial evidence demonstrates significant discrepancies between MIC and MBEC values for the same bacterial strain and antibiotic. Research on uropathogenic Escherichia coli (UPEC) biofilms reveals that while levofloxacin inhibited biofilms at 32 µg/mL, trimethoprim required concentrations between 512-1024 µg/mL to eradicate established biofilms – dramatically higher than typical planktonic MIC values [67]. This pattern extends across many bacterial species and antibiotic classes, highlighting the profound protective effect of the biofilm matrix.
Table 2: Comparative Antibiotic Efficacy Against Planktonic vs. Biofilm Cells of Uropathogenic E. coli
| Antibiotic | MIC (Planktonic Cells) | Concentration for Biofilm Inhibition/Eradication | Function |
|---|---|---|---|
| Levofloxacin | Not specified | 32 µg/mL (inhibition) | Inhibition of biofilm formation |
| Ciprofloxacin | Sub-MIC levels | Enhanced biofilm formation at sub-MIC | Paradoxical enhancement of biofilm at subinhibitory concentrations |
| Trimethoprim | Not specified | 512-1024 µg/mL (eradication) | Eradication of established biofilms |
The Protective Role of PIA in Biofilm Antibiotic Tolerance
Physical Barrier Function
The PIA-containing matrix creates a physical barrier that restricts antibiotic penetration through several mechanisms. The negatively charged PIA polymer can bind and retain certain antibiotics, while the dense matrix architecture physically impedes diffusion, creating concentration gradients where antibiotics may not reach lethal concentrations in the deeper biofilm layers [68] [69].
Alteration of Microbial Physiology
Within the PIA-encased biofilm, distinct metabolic microenvironments emerge due to nutrient and oxygen gradients. This heterogeneity results in subpopulations of slow-growing or dormant cells (persister cells) that are intrinsically less susceptible to many antibiotics that target active cellular processes [68]. Sub-minimum inhibitory concentrations of antibiotics can paradoxically enhance biofilm formation, as demonstrated with ciprofloxacin, which significantly enhanced UPEC biofilm formation at sub-MIC levels (P<0.05) [67].
Immune Evasion Mechanisms
PIA contributes significantly to immune evasion by inhibiting neutrophil phagocytosis and complement activation [3] [18]. The mechanical properties of PIA-containing biofilms also provide protection, as neutrophils can only exert stress up to approximately 1 kPa during phagocytosis, while the yield stress of mature biofilms can exceed this threshold [68]. Furthermore, neutrophils can only ingest pathogens smaller than 10 μm, thus the biofilm lifestyle protects individual bacteria and small bacterial clusters from neutrophil attack [68].
Diagram 1: PIA-mediated protection mechanisms in biofilms
Experimental Protocols for MBEC Assessment in PIA-encased Biofilms
Biofilm Cultivation and PIA Quantification
Materials Required:
- Tryptic Soy Broth (TSB) supplemented with 1% glucose
- Polystyrene microtiter plates or Calgary biofilm device pegs
- Crystal violet solution (0.1-1%) for biomass staining
- Congo Red Agar for phenotypic confirmation
- Specific enzymes (Dispersin B for PIA degradation)
Protocol:
- Inoculate bacteria in TSB with 1% glucose and adjust turbidity to 0.5 McFarland standard.
- For microtiter plate assay, add 200 μL bacterial suspension per well and incubate statically for 24-48 hours at 37°C [18].
- Remove planktonic cells by washing with phosphate-buffered saline (PBS).
- For biomass quantification, add crystal violet (0.1%) for 15 minutes, wash, and elute bound dye with 30% acetic acid. Measure absorbance at 595 nm [18].
- Confirm PIA production using Congo Red Agar plates – PIA-producing strains typically appear black with dry, crystalline consistency.
MBEC Assay Procedure
Materials Required:
- Established biofilm on peg lids or microtiter plates
- Cation-adjusted Mueller Hinton Broth
- Antibiotic serial dilutions
- Recovery media (TSB)
- ATP-based luminescence assay or viable plate counting
Protocol:
- Establish mature biofilms (typically 24-48 hours) on peg lids or in microtiter plates.
- Transfer biofilms to plates containing serial two-fold dilutions of antibiotics in cation-adjusted Mueller Hinton Broth.
- Incubate for 20-24 hours at 37°C.
- Remove antibiotic solution and wash biofilms with PBS to remove residual antibiotic.
- Transfer biofilms to recovery media containing neutralizing agents if needed.
- Determine MBEC either by:
- Viable counting: Sonicate or vortex biofilms to disaggregate cells, serially dilute, and plate on appropriate agar. MBEC is the lowest concentration reducing viable counts by ≥99.9%.
- Metabolic assays: Use resazurin, XTT, or ATP-based luminescence to measure metabolic activity post-treatment.
PIA-Specific Experimental Modifications
For PIA-specific studies, include isogenic icaADBC deletion mutants as controls. Additionally, treatment with Dispersin B (a glycosyl hydrolase that cleaves PIA) prior to antibiotic exposure can determine the specific contribution of PIA to observed tolerance [2]. Microscopy techniques including confocal laser scanning microscopy (CLSM) with PIA-specific antibodies or lectin staining can visualize matrix architecture and antibiotic penetration.
Emerging Strategies to Overcome PIA-Mediated Tolerance
Anti-PIA Immunotherapeutic Approaches
Vaccine and antibody-based strategies targeting PIA represent a promising avenue for combating biofilm infections. Antibodies against deacetylated PNAG have demonstrated protective efficacy in multiple models. Recent research shows that a combination of antibodies targeting both highly acetylated (F598 antibody) and highly deacetylated (TG10 antibody) forms of PNAG synergized to provide 90% survival in a lethal S. aureus challenge murine model [6]. This approach of "fully coating" biofilms with antibodies targeting different PIA epitopes represents a significant advance.
Enzymatic Matrix Disruption
Enzymes that degrade biofilm matrix components offer another strategic approach. Dispersin B cleaves the β-1,6-glycosidic linkages in PIA, while DNase I targets extracellular DNA, another key matrix component [2]. When used as adjuncts to conventional antibiotics, these enzymes can significantly enhance antibiotic efficacy against biofilms by improving antimicrobial penetration and disrupting the protective matrix structure.
Quorum Sensing Interference
Staphylococcal biofilm development is regulated by quorum sensing systems, which control the expression of virulence factors including PIA. Interfering with these communication systems (quorum quenching) can prevent biofilm formation or promote dispersal of established biofilms [2]. While still largely experimental, this approach represents a promising strategy for preventing biofilm-associated infections on medical devices.
Table 3: Emerging Anti-Biofilm Strategies Targeting PIA
| Strategy | Mechanism of Action | Development Stage | Key Findings |
|---|---|---|---|
| Anti-PIA/PNAG Antibodies | Opsonization, complement activation, biofilm disruption | Phase II clinical trials (F598) | Combination of F598 and TG10 antibodies provided 90% survival in murine model [6] |
| Dispersin B | Enzymatic degradation of PIA/PNAG | Preclinical research | Enhances antibiotic efficacy against staphylococcal biofilms [2] |
| dPNAG Vaccines | Induction of opsonic antibodies against deacetylated PNAG | Phase I clinical trials | Chemically deacetylated PNAG (85-100% GlcNH2) elicits protective antibodies [6] |
| Quorum Quenching | Interference with biofilm regulation | Experimental | Promotes dispersal phase; prevents biofilm formation [2] |
Diagram 2: Therapeutic strategies targeting PIA
The Scientist's Toolkit: Essential Research Reagents
Table 4: Essential Research Reagents for PIA and Biofilm Studies
| Reagent/Category | Specific Examples | Function/Application | Key Considerations |
|---|---|---|---|
| Anti-PIA Antibodies | F598, TG10, Anti-PNAG mAbs | Detection, quantification, functional blockade of PIA | F598 binds acetylated PNAG; TG10 selective for deacetylated PNAG [6] |
| Matrix-Degrading Enzymes | Dispersin B, DNase I, Proteinase K | Selective degradation of matrix components | Dispersin B specifically cleaves PIA/PNAG; useful for mechanistic studies [2] |
| Biofilm Detection Reagents | Crystal violet, Congo red, SYTO dyes | Biomass staining and quantification | Crystal violet for total biomass; fluorescent dyes for microscopy |
| Genetic Tools | icaADBC mutants, complemented strains | Establishing PIA-specific effects | Isogenic mutants essential for controlling genetic background |
| Animal Models | Murine catheter infection, prosthetic joint infection | In vivo efficacy testing | Essential for translational validation of anti-biofilm strategies |
The disconnect between MIC and MBEC values for PIA-encased staphylococcal bioframes highlights the critical limitations of current antimicrobial susceptibility testing paradigms. The PIA matrix represents both a challenge and an opportunity – its conservation across strains and species makes it an attractive target for broad-spectrum anti-biofilm strategies. Moving forward, the field must prioritize several key areas: (1) standardization of MBEC testing methodologies for clinical diagnostics; (2) development of combination therapies that target both bacterial cells and the PIA matrix; and (3) advanced understanding of PIA structural heterogeneity and its functional implications. Only through targeted approaches that acknowledge the unique biology of biofilm-embedded bacteria can we effectively address the clinical challenge of biofilm-associated infections.
Validating PIA's Role in Pathogenesis and Cross-Species Potential
Polysaccharide intercellular adhesin (PIA), also known as poly-β(1,6)-N-acetylglucosamine (PNAG), is a key exopolysaccharide component of the extracellular matrix in staphylococcal biofilms. Its role extends beyond structural support to include significant contributions to immune evasion, a critical factor in the persistence of chronic biofilm-associated infections. Staphylococcus epidermidis and Staphylococcus aureus are the primary producers of PIA, though homologous exopolysaccharides exist in other significant pathogens, including Escherichia coli (PGA) and Yersinia pestis [3]. The ability of PIA to protect encased bacteria from the host's primary cellular and chemical defenses—particularly polymorphonuclear neutrophils (PMNs) and antimicrobial peptides (AMPs)—establishes it as a central virulence determinant. This review synthesizes current mechanistic understanding of how PIA confers resistance to these immune effectors, framing it within the broader context of staphylococcal biofilm research and highlighting its implications for therapeutic development.
Structural and Biochemical Basis of PIA
PIA is a linear, positively charged polymer of β-1,6-linked N-acetylglucosamine (GlcNAc) residues with a degree of deacetylation of approximately 15-20% [3]. This partial deacetylation, resulting in free, positively charged amino groups, is critical to its function. Some N-acetylglucosamine residues (approximately 10%) may also be O-succinylated [3]. The biosynthetic machinery for PIA is encoded by the icaADBC operon, which is highly conserved in staphylococci and subject to complex environmental regulation [3].
Table 1: Key Structural Characteristics of Polysaccharide Intercellular Adhesin (PIA)
| Characteristic | Description | Functional Significance |
|---|---|---|
| Chemical Structure | Linear polymer of β-1,6-linked N-acetylglucosamine | Forms the core scaffold of the biofilm matrix [3]. |
| Molecular Weight | Variable (~30 kDa to >460 kDa reported) | Size variation may influence matrix density and integrity [3]. |
| Net Charge | Cationic (positively charged) | Crucial for electrostatic interactions with anionic surfaces and molecules [3]. |
| Deacetylation | ~15-20% of residues deacetylated | Generates positive charge; essential for resistance to antimicrobial peptides and immune evasion [3]. |
| Succinylation | ~10% of residues may be O-succinylated | May modulate the overall charge and physical properties of the polymer [3]. |
PIA-mediated Protection against Polymorphonuclear Neutrophils (PMNs)
PMNs are the body's first line of cellular defense against bacterial infections, employing mechanisms such as phagocytosis, the release of reactive oxygen species (ROS), and the formation of neutrophil extracellular traps (NETs). PIA enhances biofilm resilience through multiple mechanisms that impair these neutrophil functions.
Physical Barrier and Inhibition of Phagocytosis
The PIA-containing extracellular matrix acts as a physical barrier that limits neutrophil penetration and access to bacterial cells. Confocal microscopy studies have demonstrated that immune cells, including macrophages and neutrophils, cannot effectively infiltrate intact biofilms or phagocytose biofilm-associated bacteria [70] [71]. The matrix's physical structure is a primary factor in this exclusion. Furthermore, the presence of extracellular DNA (eDNA) within the biofilm matrix, which often co-localizes with PIA, adds another layer of protection. Specific Z-form DNA (Z-DNA) has been identified as a major structural component in staphylococcal biofilms, contributing to resistance against phagocytosis by PMNs [70].
Electrostatic Shielding and Resistance to Opsonization
The cationic nature of PIA, conferred by its deacetylated amino groups, facilitates electrostatic interactions with anionic molecules on host cell surfaces. This property can interfere with the opsonization process, a critical step for efficient phagocytosis. Antibodies generated against PIA have been shown to exhibit opsonic activity. One study reported that anti-PIA antibodies induced an opsonophagocytic killing effect of 40.48% against E. coli, demonstrating that targeting PIA can enhance bacterial clearance by professional phagocytes [18]. This suggests that native PIA may normally shield bacterial surface antigens from recognition by opsonizing antibodies.
Table 2: Quantitative Data on PIA-mediated Immune Evasion and Targeting
| Immune Effector / Intervention | Experimental Finding | Significance / Implication |
|---|---|---|
| Anti-PIA Antibodies (Opsonization) | 40.48% lethality in E. coli via opsonophagocytic killing [18] | Demonstrates PIA is a viable target for protective antibodies. |
| Anti-PIA Antibodies (Biofilm Inhibition) | Significant inhibition of in vitro biofilm formation in E. coli ATCC 25922 [18] | Antibodies can disrupt the biofilm matrix integrity. |
| Macrophage Infiltration | Increased invasion and attachment to ΔsaeRS and ΔfnbA mutant biofilms [70] | SaeRS-regulated factors (e.g., FnbA) contribute to the physical barrier. |
| Biofilm Structure | Δeap (eDNA-binding protein) biofilms show altered structure and increased macrophage phagocytosis [70] | eDNA and associated proteins are critical for biofilm barrier function. |
The following diagram illustrates the primary mechanisms by which PIA protects bacterial biofilms from neutrophil attacks:
PIA-mediated Resistance to Antimicrobial Peptides (AMPs)
AMPs, such as defensins and cathelicidins, are cationic peptides that disrupt bacterial membranes. A key mechanism of PIA-mediated resistance involves electrostatic interaction with these host defense molecules.
Cationic Sequestration and Neutralization
The cationic character of PIA, arising from its deacetylated residues, allows it to function as a molecular "sponge" for cationic AMPs [3] [72]. The negative charges on bacterial membranes (e.g., from teichoic acids) and the lipid A moiety of lipopolysaccharide attract cationic AMPs. By presenting a high density of positive charges in the extracellular space, PIA can electrostatically bind and sequester AMPs, preventing them from reaching and disrupting the bacterial cytoplasmic membrane [72]. This neutralization is a primary mechanism for AMP resistance in staphylococcal biofilms.
Experimental Methodologies for Investigating PIA Function
Studying PIA's role in immune evasion requires a combination of microbiological, biochemical, and immunological techniques. Below are detailed protocols for key experiments.
PIA Extraction and Purification Protocol
The following method, adapted from established procedures, is used to isolate PIA from Staphylococcus epidermidis [18]:
- Bacterial Culture: Inoculate S. epidermidis (e.g., strain 1457) into 2 liters of Tryptic Soy Broth (TSB). Incubate at 37°C for 24 hours with gentle agitation (40–50 rpm).
- Harvesting and Washing: Centrifuge the culture at 1,000 × g for 20 minutes at 4°C. Discard the supernatant and resuspend the bacterial pellet in 20 mL of phosphate-buffered saline (PBS), pH 7.5.
- Cell Disruption: Sonicate the cell suspension on ice using four cycles of 30 seconds each.
- Debris Removal: Centrifuge the sonicated sample at 12,000 rpm for 15 minutes to remove cell debris. Collect the supernatant, which contains the crude polysaccharide extract.
- Enzymatic Treatment: Treat the supernatant with DNase and RNase (e.g., 20 µg/mL) for 1 hour at 37°C, followed by proteinase K (e.g., 100 µg/mL) for an additional hour at 37°C to digest nucleic acids and proteins.
- Dialysis and Precipitation: Dialyze the treated supernatant extensively against distilled water. Precipitate the polysaccharide by adding trichloroacetic acid (TCA) to a final concentration of 5-10% (w/v) or by adding 3-4 volumes of cold ethanol.
- Purification: Recover the precipitate by centrifugation, re-dissolve it in a small volume of PBS or water, and purify further using size-exclusion chromatography (e.g., Sephadex G-200 column).
- Confirmation: Confirm the identity and purity of the extracted PIA using Fourier-Transform Infrared Spectroscopy (FTIR), Nuclear Magnetic Resonance (NMR), and colorimetric assays for hexosamine content [18].
In Vitro Biofilm Formation and Inhibition Assay
This semi-quantitative method assesses biofilm formation and the inhibitory effect of anti-PIA antibodies [18]:
- Preparation of Inoculum: Grow the test strain (e.g., E. coli ATCC 25922) in Brain Heart Infusion (BHI) broth. Adjust the optical density of the culture to OD₆₀₀ = 0.7.
- Dilution and Supplementation: Dilute the bacterial culture 1:200 in BHI broth supplemented with 1% (w/v) D-glucose (BHIGlc) and 4% (w/v) NaCl (BHINaCl) to induce robust biofilm formation.
- Incubation: Transfer 200 µL of the diluted culture into the wells of a sterile polystyrene microtiter plate (e.g., Nunc). Incubate the plate for 24 hours at 37°C under static conditions to allow biofilm adhesion.
- Staining: After incubation, carefully wash each well three times with PBS to remove non-adherent planktonic cells. Add 150 µL of a 1% crystal violet solution to each well and stain for 15-30 minutes.
- Destaining and Quantification: Wash the wells again to remove excess stain. Elute the crystal violet bound to the adherent cells by adding 160 µL of 30% acetic acid to each well.
- Measurement: Measure the absorbance of the eluted dye spectrophotometrically at 595 nm. Higher absorbance values correlate with greater biofilm biomass. To test antibody inhibition, include wells containing bacteria co-cultured with anti-PIA serum or purified anti-PIA antibodies and compare absorbance to controls (e.g., non-immune serum).
Opsonophagocytic Killing Assay
This functional assay quantifies the ability of anti-PIA antibodies to promote bacterial killing by phagocytes [18]:
- Preparation of Components:
- Bacteria: Harvest log-phase bacteria, wash, and resuspend in PBS.
- Phagocytes: Isolate human or murine PMNs from fresh blood using density gradient centrifugation.
- Complement: Use fresh serum from a non-immune host as a complement source (or complement-deficient serum as a control).
- Antibody: Purified anti-PIA antiserum or monoclonal antibodies.
- Reaction Mixture: In a microcentrifuge tube, combine the following:
- Bacteria (e.g., ~1 × 10⁴ CFU)
- Phagocytes (e.g., at an effector-to-target ratio of 50:1 or 100:1)
- Anti-PIA antibody (at a predetermined concentration)
- Complement source (e.g., 5-10% final concentration)
- Complete the volume with tissue culture medium containing opsonophagocytosis buffer.
- Incubation and Control Setup: Incubate the reaction mixture with rotation or end-over-end mixing for 60-90 minutes at 37°C. Include control reactions lacking phagocytes, antibody, or complement to determine baseline survival.
- Viability Assessment: After incubation, serially dilute the reactions in sterile water or PBS to lyse the phagocytes. Plate the dilutions onto agar plates and incubate overnight at 37°C.
- Calculation: Count the colony-forming units (CFU) the next day. Calculate the percentage of bacterial killing using the formula: % Killing = [1 - (CFU from test reaction / CFU from control reaction without antibody)] × 100
The following workflow summarizes the key experimental approaches for studying PIA:
The Scientist's Toolkit: Key Research Reagents
Table 3: Essential Reagents for Investigating PIA-mediated Immune Evasion
| Reagent / Material | Function / Application | Example Usage |
|---|---|---|
| S. epidermidis 1457 | A standard, well-characterized PIA-producing strain. | Positive control for PIA extraction, biofilm formation, and genetic studies [18]. |
| Δica Mutant Strains | Isogenic mutants with a disrupted PIA biosynthetic operon. | Essential negative control to confirm PIA-specific phenotypes [18]. |
| Tryptic Soy Broth (TSB) | Nutrient-rich growth medium for staphylococci. | Routine culture and induction of PIA/biofilm formation, especially under static, low-agitation conditions [18]. |
| Polyclonal Anti-PIA Antiserum | Antibodies targeting PIA/PNAG. | Used in opsonophagocytic killing assays, biofilm inhibition studies, and immuno-detection (e.g., ELISA, microscopy) [18]. |
| Polymorphonuclear Neutrophils (PMNs) | Primary human or murine phagocytic cells. | Effector cells in functional opsonophagocytic killing assays to measure PIA-dependent immune evasion [18] [71]. |
| Complement Source | Active serum from a non-immune animal (e.g., baby rabbit). | Critical component in opsonophagocytosis assays to complement the action of anti-PIA antibodies [18]. |
| Crystal Violet | A simple histological dye that binds to negatively charged surface molecules. | Semi-quantitative staining of adherent biofilm biomass in microtiter plate assays [18]. |
| DNase, RNase, Proteinase K | Enzymes for digesting nucleic acids and proteins. | Used during PIA extraction to purify the polysaccharide from other matrix components (eDNA, proteins) [18]. |
PIA is a multifunctional virulence factor that underpins biofilm-associated chronic infections by providing a robust defense against two pivotal arms of the innate immune system: phagocytic cells (like PMNs) and antimicrobial peptides. Its mechanisms are twofold, acting as a physical barrier that impedes cellular penetration and phagocytosis, and as an electrostatic shield that neutralizes cationic antimicrobial peptides. The experimental data demonstrating that antibodies targeting PIA can disrupt biofilms and promote opsonophagocytic killing, with a reported lethality of 40.48% [18], underscore its potential as a therapeutic target. Future research should focus on translating this understanding into clinical applications, such as the development of PIA-targeting vaccines or monoclonal antibodies, which could be used as adjuvants to conventional antibiotics to clear recalcitrant biofilm infections.
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), is a key exopolysaccharide component of the staphylococcal biofilm matrix. Its production, mediated by the icaADBC operon, has been epidemiologically correlated with device-related infections, particularly those involving Staphylococcus epidermidis and Staphylococcus aureus. This whitepaper synthesizes clinical and experimental evidence establishing PIA as a major virulence determinant in implant-associated infections. We summarize key epidemiological data, detail the molecular mechanisms underpinning its role in pathogenesis, and present standardized methodologies for its investigation. The collective evidence positions PIA as a critical target for novel diagnostic and therapeutic strategies against biofilm-related infections.
Polysaccharide intercellular adhesin (PIA) is a linear, cationic exopolysaccharide composed of β-1-6-linked N-acetylglucosamine residues that are partially deacetylated (approximately 15-20%) [3]. This chemical structure confers positive charges that are crucial for its function. PIA serves as the primary structural scaffold for the extracellular matrix of staphylococcal biofilms, facilitating both cell-to-cell adhesion and surface attachment [1]. The biosynthetic pathway for PIA is encoded by the icaADBC (intercellular adhesion) operon, with the IcaA, IcaD, IcaB, and IcaC proteins each playing distinct roles in its synthesis, deacetylation, and export [3] [1]. Beyond its structural role, PIA provides a physical barrier that significantly enhances bacterial resistance to host innate immune defenses, including phagocytosis and antimicrobial peptides, thereby promoting persistent infections [53] [73]. The epidemiological significance of PIA stems from its strong association with infections involving indwelling medical devices, such as prosthetic joints, central venous catheters, and other implanted materials [3] [73].
Epidemiological and Clinical Evidence Linking PIA to Device-Related Infections
Epidemiological studies consistently demonstrate a higher prevalence of the ica locus and PIA production in clinical isolates obtained from device-associated infections compared to commensal isolates. The table below summarizes the key epidemiological findings:
Table 1: Epidemiological Evidence for PIA in Device-Related Infections
| Evidence Type | Key Finding | Significance | Reference |
|---|---|---|---|
| Genetic Association | The ica genes are virtually the only genes in S. epidermidis with a significantly higher prevalence in device-infection isolates versus colonization isolates. | Strongly links the ica locus to pathogenic infection rather than harmless colonization. | [3] |
| Species-Specific Clustering | In S. epidermidis, ica genes are predominantly found in a specific genetic cluster (A/B), which is associated with infectivity. | Suggests a population-level genetic basis for PIA-associated virulence. | [3] |
| Causative Agent Prevalence | S. epidermidis and S. aureus, both PIA producers, account for 30-43% and 12-23% of prosthetic joint infections (PJIs), respectively. | Establishes PIA-producing species as the leading etiologic agents of device-related infection. | [73] |
| Functional Validation | Isogenic mutants lacking PIA production (e.g., 1457-M10) show increased susceptibility to phagocytosis and killing in ex vivo and in vivo models. | Confirms a direct, functional role for PIA in immune evasion and pathogen survival. | [53] [73] |
| Immune Response Correlation | High levels of complement C5a are detected in sera from PJI patients infected with PIA-producing S. epidermidis strains. | Indicates that PIA production triggers a specific and measurable host immune response during clinical infection. | [73] |
While the association is strong, some studies have questioned a universal correlation, indicating that alternative, ica-independent biofilm mechanisms exist in some strains [3] [1]. However, the collective epidemiological evidence firmly establishes PIA as a major risk factor for the development of chronic, device-associated staphylococcal infections.
Molecular Mechanisms of PIA in Pathogenesis
PIA contributes to device-related pathogenesis through two primary, interconnected mechanisms: biofilm formation and immune evasion.
Biofilm Formation and Architecture
PIA is the fundamental mediator of biofilm accumulation in staphylococci. It enables the formation of a structured, multi-layered biofilm on the surface of implanted medical devices. This biofilm architecture creates a protected niche for bacterial proliferation, reduces metabolic activity, and acts as a diffusion barrier, dramatically increasing tolerance to conventional antibiotics [3] [1].
Immune Evasion
PIA provides a robust defense against the human innate immune system. Key mechanisms include:
- Inhibition of Phagocytosis: PIA forms fibrous strands on the bacterial surface that significantly reduce phagocytosis and killing by human polymorphonuclear leukocytes (PMNs) [53].
- Resistance to Antimicrobial Peptides: PIA protects staphylococci from major cationic antibacterial peptides of human skin, such as human β-defensin 3 and LL-37, as well as the anionic peptide dermcidin [53].
- Modulation of Complement Activation: The PIA-dependent biofilm matrix inhibits the deposition of opsonic complement components (e.g., C3b) and immunoglobulin G (IgG) on the bacterial surface. Concurrently, it can induce the production of the immunosuppressive complement component C5a, which further impairs neutrophil function [73].
Phase Variation and Regulation
The expression of PIA is a dynamic process, strongly influenced by environmental conditions and subject to complex regulation. A notable mechanism of regulation is phase variation. In S. aureus, reversible, RecA-independent expansion and contraction of a tetranucleotide tandem repeat within the icaC gene can lead to a frameshift mutation, effectively turning PIA production on and off [25]. This phase variation allows a bacterial population to maintain heterogeneity, potentially enabling immune evasion by subpopulations that cease production of this dominant antigen [25].
Experimental Protocols for PIA Research
To ensure reproducibility and standardization across studies, key experimental protocols for PIA investigation are detailed below.
PIA Extraction and Purification Protocol
This protocol is adapted from established methods for extracting native PIA from staphylococcal cultures [18] [31].
- Culture and Harvest: Inoculate S. epidermidis (e.g., strain 1457) into 2 liters of Tryptic Soy Broth (TSB). Incubate at 37°C for 24 hours with gentle agitation (40-50 rpm). Harvest cells by centrifugation (e.g., 4,500 rpm for 20 minutes at 4°C).
- Cell Lysis: Resuspend the cell pellet in 20 mL of phosphate-buffered saline (PBS, pH 7.5). Subject the suspension to sonication on ice (four cycles of 30 seconds each).
- Clarification and Digestion: Centrifuge the lysate at 12,000 rpm for 15 minutes at 4°C to remove cell debris. Transfer the supernatant to a new tube and treat with proteinase K to degrade soluble proteins.
- Dialysis and Concentration: Dialyze the supernatant overnight against PBS using a 12 kDa molecular weight cut-off membrane. Concentrate the retentate using centrifugal concentrators (e.g., Centriprep 10, Amicon).
- Size-Exclusion Chromatography: Load the concentrated sample onto a Sephacryl S-100 column (e.g., 1.6 x 100 cm) pre-equilibrated with 50 mM sodium phosphate buffer. Elute at a flow rate of 1 mL/min, collecting 5 mL fractions.
- Analysis and Storage: Analyze fractions for hexosamine content using colorimetric assays (e.g., MBTH assay). Pool positive fractions, confirm structure via FTIR or NMR, and store purified PIA at -20°C [31].
In Vitro Biofilm Formation and Inhibition Assay
This semi-quantitative method is widely used to assess biofilm formation and the efficacy of inhibitory agents like antibodies [18] [31].
- Preparation: Adjust the optical density (OD600) of an overnight bacterial culture to 0.7. Dilute the culture 1:200 in Brain Heart Infusion (BHI) broth supplemented with 1% glucose (BHIGlc).
- Incubation: Transfer 200 µL of the diluted suspension into the wells of a polystyrene microtiter plate. For inhibition assays, include test substances (e.g., anti-PIA antiserum) in the wells. Include control wells with sterile broth (negative control) and bacteria without inhibitor (positive control).
- Growth and Staining: Incubate the plate for 24 hours at 37°C. Carefully remove planktonic cells and wash each well three times with PBS to remove non-adherent bacteria.
- Fixation and Staining: Add 150 µL of a 1% crystal violet solution to each well and incubate for several minutes. Discard the stain and wash the wells thoroughly with water to remove excess dye.
- Elution and Quantification: Dissolve the crystal violet bound to the adherent biofilm in 160 µL of 30% acetic acid. Measure the absorbance of the eluted dye spectrophotometrically at 595 nm.
- Calculation of Inhibition: Calculate the percentage of biofilm inhibition using the formula:
% Inhibition = [(A595_positive_control - A595_antibody) / (A595_positive_control - A595_negative_control)] * 100[31].
The Scientist's Toolkit: Key Research Reagents
The following table catalogues essential reagents and their applications in PIA and biofilm research, as derived from the cited methodologies.
Table 2: Key Research Reagents for PIA and Biofilm Studies
| Reagent / Material | Function in Research | Example Application |
|---|---|---|
| S. epidermidis 1457 | A well-characterized model strain that produces a PIA-dependent biofilm. | Used as a standard positive control for biofilm formation and PIA extraction studies [73]. |
| Isogenic Mutant (e.g., 1457-M10) | A mutant derived from strain 1457 with a disrupted icaADBC locus; does not produce PIA/biofilm. | Serves as a critical negative control to isolate the specific effects of PIA in comparative experiments [73]. |
| Sephacryl S-100 Resin | A matrix for size-exclusion chromatography. | Used for the final purification step of native PIA polysaccharide from crude cell extracts [31]. |
| Anti-PIA/PNAG Antibodies | Polyclonal or monoclonal antibodies raised against the purified polysaccharide. | Used to probe the role of PIA in immune evasion, opsonic killing, and biofilm inhibition in vitro and in vivo [18] [31]. |
| Congo Red Agar (CRA) | A differential growth medium. | Used to visually screen for PIA-producing colonies based on their dry, black morphologies versus the slick, red colonies of non-producers [25]. |
| Polystyrene Microtiter Plate | A non-treated, sterile surface for biofilm growth. | The standard vessel for high-throughput, semi-quantitative biofilm assays using crystal violet staining [18]. |
The epidemiological evidence firmly establishes PIA as a major virulence factor in staphylococcal device-related infections. Its dual role in mediating robust biofilm formation on inert surfaces and protecting embedded bacteria from host immune defenses makes it a critical contributor to the chronic and recalcitrant nature of these infections. The experimental frameworks and reagents outlined herein provide a foundation for ongoing research.
Future efforts are increasingly focused on leveraging this knowledge to develop novel therapeutic and preventive strategies. The high conservation of PIA across staphylococcal species and its surface accessibility make it an attractive target for immunoprophylaxis. Promising approaches include the development of vaccines based on the deacetylated form of PNAG to enhance opsonophagocytic killing [1] and the use of monoclonal antibodies to disrupt biofilm integrity and sensitize bacteria to conventional antibiotics [18] [31]. Validating these interventions in clinical settings represents the next critical step in translating our epidemiological and molecular understanding of PIA into improved patient outcomes.
Abstract Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), is a key constituent of the extracellular matrix in staphylococcal biofilms. Its production is mediated by the icaADBC operon. This whitepaper provides a comparative analysis of the virulence of PIA-positive versus PIA-negative isogenic mutants of Staphylococcus epidermidis and Staphylococcus aureus, the leading etiologic agents of biomaterial-associated infections. Framed within the broader context of PIA's role in biofilm research, this guide synthesizes data on the molecular function of PIA, its critical role in immune evasion, and its contribution to in vivo pathogenicity. Structured quantitative data, detailed experimental protocols for key assays, and essential research tools are provided to support drug development professionals in designing targeted anti-biofilm strategies.
The success of staphylococci as pathogens in device-related infections is intrinsically linked to their ability to form biofilms. A key component of the staphylococcal biofilm matrix is polysaccharide intercellular adhesin (PIA), a linear, positively charged polymer of β-1-6-linked N-acetylglucosamine that is partially deacetylated (approximately 15-20%) [3] [1]. The biosynthesis of PIA is encoded by the icaADBC operon, with the IcaA and IcaD proteins synthesizing the nascent polysaccharide chain, IcaC facilitating its export, and IcaB responsible for the crucial deacetylation that imparts a positive charge [2] [11]. The use of isogenic mutants—genetically identical strains differing only in the presence or absence of a functional ica locus—has been instrumental in isolating and quantifying the specific contribution of PIA to virulence, separate from other bacterial factors [73].
Molecular and Functional Basis of PIA-Mediated Virulence
PIA's structure is fundamental to its function. The deacetylation of N-acetylglucosamine residues creates a cationic polymer, which enables electrostatic interactions with anionic bacterial cell surfaces and other matrix components [3] [11]. This is critical for intercellular adhesion and the structural integrity of the biofilm. Beyond its role as a mechanical scaffold, PIA is a potent modulator of the host immune response.
- Intercellular Adhesion and Biofilm Architecture: PIA is the primary mediator of the accumulation phase of biofilm formation, enabling the formation of multi-layered, structured communities [62]. PIA-negative mutants often form biofilms with significantly reduced biomass and structural stability.
- Immune Evasion: PIA confers resistance to key host defense mechanisms. It has been demonstrated to inhibit the deposition of complement factors and immunoglobulin G (IgG) on the bacterial surface, thereby reducing opsonophagocytosis by neutrophils [73] [71]. Furthermore, PIA can confer resistance to antimicrobial peptides [3].
Quantitative Comparison of Virulence Phenotypes
The following tables summarize key phenotypic differences between PIA-positive and PIA-negative isogenic mutants, as established in experimental models.
Table 1: In Vitro Phenotypic Comparisons
| Virulence Phenotype | PIA-Positive Strain | PIA-Negative Isogenic Mutant | Key Experimental Findings & Context |
|---|---|---|---|
| Biofilm Formation | Robust, multi-layered biofilm [62] | Significantly impaired or absent biofilm accumulation [73] [62] | Quantified using microtiter plate assays and scanning electron microscopy. |
| Resistance to Phagocytosis | High resistance to uptake and killing by polymorphonuclear neutrophils (PMNs) [71] | Significantly more susceptible to phagocytosis and killing by PMNs [71] | Assessed via in vitro phagocytosis assays using isolated human PMNs. |
| Complement Activation | Induces high levels of complement component C5a [73] | Induces lower levels of C5a [73] | Measured in an ex vivo human whole blood infection model. |
| Antimicrobial Peptide (AMP) Resistance | Enhanced resistance to AMPs [3] [11] | Increased susceptibility to AMPs [3] | Determined by minimum inhibitory concentration (MIC) assays. |
Table 2: In Vivo Virulence in Animal Models
| Infection Model | PIA-Positive Strain | PIA-Negative Isogenic Mutant | Interpretation and Significance |
|---|---|---|---|
| Biomaterial-Associated Infection (e.g., Catheter, Prosthetic Joint) | Persistent infection; high bacterial load recovered from the implant [73] [62] | Significantly reduced bacterial load; infection often cleared [73] [62] | Demonstrates that PIA is critical for establishing chronic, device-related infections. |
| Caenorhabditis elegans Killing | Lethal to nematodes [73] | Attenuated killing ability [73] | Used as a model for innate immune evasion and virulence. |
Essential Experimental Protocols
To generate the comparative data outlined above, specific, reproducible methodologies are required. Below are detailed protocols for two fundamental assays.
Protocol: Ex Vivo Whole Blood Killing and Immune Activation Assay
This assay evaluates bacterial survival in human blood and the subsequent immune response, particularly complement activation [73].
- Bacterial Preparation: Grow the PIA-positive (e.g., S. epidermidis 1457) and PIA-negative isogenic mutant (e.g., S. epidermidis 1457-M10) strains to stationary phase in a suitable broth like Tryptic Soy Broth (TSB). Harvest, wash, and resuspend the bacteria in phosphate-buffered saline (PBS). Adjust the optical density to prepare defined inocula (e.g., 10^4 - 10^7 CFU/mL).
- Blood Infection: Draw fresh, heparinized human whole blood from healthy volunteers. Add 100 µL of the bacterial suspension to 900 µL of whole blood in a sterile tube.
- Incubation: Incubate the blood-bacteria mixture at 37°C with constant rotation or gentle shaking for up to 24 hours.
- Sample Analysis:
- Bacterial Killing: At designated timepoints (e.g., 0, 3, 6, 24h), remove aliquots, perform serial dilutions in PBS, and plate on agar for Colony Forming Unit (CFU) enumeration after overnight incubation.
- Complement C5a Measurement: Centrifuge blood samples at the end of the incubation to collect plasma. Use a commercial enzyme-linked immunosorbent assay (ELISA) kit specific for human C5a to quantify its concentration in the plasma.
Protocol: In Vitro Biofilm Formation Assay (Microtiter Plate Method)
This is a standard, semi-quantitative method to assess biofilm-forming capacity [18].
- Inoculum Preparation: Grow test strains overnight in TSB. Dilute the culture 1:200 in a biofilm-inducing medium, such as TSB supplemented with 1% (w/v) glucose (TSBG) or Brain Heart Infusion (BHI) broth with 1% glucose and 4% NaCl (BHINaCl).
- Biofilm Growth: Dispense 200 µL of the diluted culture into the wells of a sterile, flat-bottomed polystyrene microtiter plate. Include wells with sterile broth as negative controls. Incubate the plate statically for 24 hours at 37°C.
- Biofilm Staining and Quantification:
- Carefully remove the planktonic cells and medium by inverting and shaking the plate. Wash the adhered biofilms three times with PBS to remove non-adherent cells.
- Fix the biofilms by air-drying or using methanol.
- Stain the biofilms with 150 µL of a 1% (w/v) crystal violet solution for 15 minutes.
- Rinse the plate thoroughly under running tap water to remove excess stain.
- Elute the bound crystal violet by adding 160 µL of 30% (v/v) acetic acid to each well.
- Transfer 125 µL of the eluent to a new microtiter plate and measure the optical density (OD) at 595 nm using a microplate reader. The OD value is proportional to the amount of biofilm formed.
Visualization of PIA Biosynthesis and Experimental Workflow
Diagram 1: PIA Biosynthesis and Virulence Pathway. This diagram outlines the genetic regulation and enzymatic machinery of PIA production and its role in virulence.
Diagram 2: Experimental Workflow for Virulence Comparison. This flowchart illustrates the parallel evaluation of PIA-positive and PIA-negative mutants across key functional assays.
The Scientist's Toolkit: Key Research Reagents and Models
Table 3: Essential Research Tools for PIA Virulence Studies
| Reagent / Model | Function and Application | Example/Description |
|---|---|---|
| Isogenic Mutant Pair | Gold standard for isolating PIA's specific role by controlling for genetic background. | S. epidermidis 1457 (PIA+) / 1457-M10 (PIA-, icaADBC::Tn) [73]. |
| Anti-PIA/PNAG Antibodies | Used for immunological detection, opsonophagocytosis assays, and as a potential therapeutic. | Rabbit or mouse IgG targeting deacetylated PNAG; used to confirm PIA expression and function [18]. |
| Dispersin B | A glycoside hydrolase that cleaves PIA/PNAG; used as a tool to chemically validate PIA's role. | Enzyme from Aggregatibacter actinomycetemcomitans; applied to degrade pre-formed biofilms [2]. |
| Proteases & DNases | Used to determine the composition of the biofilm matrix and identify PIA-independent mechanisms. | Proteinase K and DNase I are used to treat biofilms in parallel with PIA-targeting tools [2] [11]. |
| Ex Vivo Whole Blood Model | Models human innate immune response, including phagocytosis and complement activation. | Fresh heparinized human blood infected with bacteria to assess survival and immune activation (C5a) [73]. |
| Prosthetic Joint Infection (PJI) Model | In vivo model reflecting the clinical context of device-related biofilm infections. | Rodent model where a foreign material is implanted in the joint and infected with staphylococci [73]. |
The comparative analysis of PIA-positive and PIA-negative isogenic mutants unequivocally establishes PIA as a master virulence factor in staphylococcal biofilm-associated infections. Its role extends beyond physical cohesion to active interference with host immunity, making it a compelling target for novel therapeutic and prophylactic strategies. Current research focuses on PIA as a vaccine component, the development of PIA-degrading enzymes like Dispersin B, and the exploration of quorum-sensing inhibitors to downregulate ica expression [3] [2]. Future work must further elucidate the complex regulatory network controlling PIA production and its interaction with other matrix components in mixed-species biofilms, paving the way for next-generation anti-biofilm agents to combat chronic, device-related infections.
Polysaccharide Intercellular Adhesin (PIA), a key biofilm matrix component in staphylococci, shares striking biochemical similarity with poly-β(1,6)-N-acetylglucosamine (PNAG) produced by other pathogens, including Escherichia coli. This structural conservation presents a unique opportunity for cross-reactive immunity. Recent research demonstrates that antibodies generated against Staphylococcus epidermidis-derived PIA exhibit significant functional activity against E. coli biofilms, reducing biofilm formation and enhancing bacterial opsonization. This whitepaper synthesizes evidence from foundational and cutting-edge studies to provide a technical guide on the mechanisms, experimental validation, and therapeutic potential of anti-PIA antibodies in targeting phylogenetically distinct bacterial biofilms, framing these findings within the broader context of PIA research and anti-biofilm drug development.
Polysaccharide Intercellular Adhesin (PIA) is a paramount virulence factor in staphylococcal biofilm research. In Staphylococcus epidermidis and Staphylococcus aureus, PIA is a cationic exopolysaccharide that forms the structural backbone of the biofilm matrix, enabling cell-to-cell adhesion, community stability, and protection against host immune defenses [1] [2]. Its structure is defined as a poly-β(1-6)-N-acetylglucosamine (PNAG), partially deacetylated, which contributes to its positive charge and adhesive properties [1] [2]. The synthesis of PIA is mediated by the icaADBC (intercellular adhesion) operon. The product of icaA is an N-acetylglucosaminyltransferase that synthesizes PIA oligomers, icaD enhances the efficiency of IcaA, icaC is involved in the externalization of the polysaccharide, and icaB encodes an N-deacetylase responsible for the partial deacetylation of the polymer [1] [2]. The deacetylation generates a positive charge, crucial for the intercellular adhesive properties and for interaction with negatively charged host molecules [2].
The role of PIA extends beyond mere structural support; it is a critical factor in the pathogenesis of implant-associated infections. PIA provides protection against opsonophagocytosis and the activity of antimicrobial peptides, thereby facilitating immune evasion and chronicity of infections [74]. Consequently, PIA has been a compelling target for opsonic and protective antibodies, as well as a candidate for vaccine development against staphylococcal infections [1] [75]. The discovery that other bacteria, including E. coli, produce a biochemically indistinguishable exopolysaccharide, also termed PNAG, has expanded the potential scope of PIA-targeted immunological strategies, suggesting the possibility of cross-protective immunity across different bacterial genera [18].
Structural and Functional Homology Between Staphylococcal PIA and E. coli PNAG
The foundation for cross-reactive immunity lies in the profound structural and functional homology between staphylococcal PIA and E. coli PNAG.
- Biochemical Identity: PIA in staphylococci and the exopolysaccharide produced by E. coli are biochemically indistinguishable; both are forms of poly-β(1,6)-N-acetylglucosamine [18] [75]. This shared structure is the molecular basis for the cross-reactivity observed with antibodies and other binding molecules.
- Functional Convergence: In both organisms, this polysaccharide is a major functional factor in biofilm formation. It promotes cell-to-cell aggregation, stabilizes the three-dimensional architecture of the biofilm matrix, and provides protection against environmental stresses and host immune defenses [18]. In E. coli, the synthesis is mediated by the pgaABCD operon, which is functionally analogous to the staphylococcal icaADBC operon [18].
This homology indicates an evolutionary convergence on a highly effective polysaccharide for building resilient microbial communities, which can be exploited therapeutically by targeting this common molecule.
Experimental Evidence of Cross-Reactive Effects
A pivotal 2025 study directly investigated the effect of anti-PIA antibodies on E. coli biofilm formation and opsonization [18]. The key experimental findings are consolidated in the table below.
Table 1: Summary of Key Experimental Findings on Cross-Reactive Anti-PIA Antibodies
| Experimental Parameter | Finding | Significance |
|---|---|---|
| PIA Purification Source | Staphylococcus epidermidis | Confirmed using FTIR, NMR, and colorimetric methods [18]. |
| Animal Model for Antibody Generation | Mouse model | Immunized with purified S. epidermidis PIA [18]. |
| Antibody Response | Significant increase in IgG titers in immunized vs. control groups | Demonstrated robust immunogenicity of the purified PIA antigen [18]. |
| In Vitro Biofilm Inhibition | Significant difference in inhibiting E. coli (ATCC 25922) biofilm production | Anti-PIA antibodies directly disrupted biofilm integrity [18]. |
| Opsonic Killing Activity | 40.48% lethality against E. coli | Antibodies facilitated immune-mediated clearance of the bacteria [18]. |
| In Vivo Lethality (Mouse Mastitis Model with S. aureus) | Strong biofilm-forming strain advanced time of mortality | Suggests biofilm influences virulence and disease progression, underscoring the need for effective countermeasures [76]. |
This data provides quantitative evidence that antibodies targeting a staphylococcal antigen can confer functional, cross-reactive effects against a Gram-negative pathogen, E. coli.
Detailed Experimental Protocols
This section details the core methodologies used in the cited research to evaluate anti-PIA antibodies and their cross-reactive effects.
PIA Extraction and Purification from Staphylococcus epidermidis
The following protocol, adapted from established methods, is used for PIA extraction and purification [18] [31]:
- Culture and Harvest: Inoculate S. epidermidis (e.g., strain 1457) in 2 liters of Tryptic Soy Broth (TSB). Incubate at 37°C for 24 hours with gentle agitation (40–50 rpm). Harvest bacterial cells by centrifugation at 4°C [18].
- Cell Lysis and Primary Purification: Resuspend the cell pellet in phosphate-buffered saline (PBS), pH 7.5. Sonicate the suspension on ice. Centrifuge to remove cell debris. The supernatant is dialyzed against PBS and concentrated using centrifugal filter devices (e.g., Centriprep 10, Amicon) [31].
- Enzymatic Digestion and Chromatography: Treat the concentrated supernatant with proteinase K to eliminate contaminating proteins. Load the sample onto a size-exclusion chromatography column (e.g., Sephacryl S-100 or Sephacryl S-300) equilibrated with 50 mM sodium phosphate buffer. Elute fractions and monitor for polysaccharide content [31].
- Validation: Confirm the identity and purity of the extracted PIA using Fourier-Transform Infrared Spectroscopy (FTIR), Nuclear Magnetic Resonance (NMR), and colorimetric assays for hexosamine content (e.g., MBTH method) [18] [31].
Generation and Evaluation of Anti-PIA Antibodies
- Animal Immunization: Female BALB/c mice are subcutaneously immunized three times at two-week intervals (e.g., days 0, 14, and 28) with 100 µg of purified PIA adsorbed to 1% alum adjuvant. Control groups receive PBS or adjuvant alone [31].
- Serum Collection and Analysis: Collect blood samples two weeks after each immunization. Isolate serum and evaluate the anti-PIA antibody titer using an enzyme-linked immunosorbent assay (ELISA). Coat ELISA plates with purified PIA, apply serial dilutions of mouse serum, and detect bound IgG with enzyme-conjugated anti-mouse IgG antibodies [18] [31].
In Vitro Biofilm Inhibition Assay
A standard semi-quantitative method is used to assess the impact of antibodies on biofilm formation [18] [31]:
- Biofilm Growth: Prepare a bacterial suspension (e.g., E. coli ATCC 25922) in a growth medium such as Brain Heart Infusion broth supplemented with 1% glucose. Dilute the suspension and add it to a polystyrene microtiter plate. Include test wells containing a pre-determined dilution of anti-PIA serum or purified IgG. Wells with non-immune serum or PBS serve as negative controls.
- Incubation and Staining: Incub the plate under static conditions for 24 hours at 37°C. After incubation, gently wash the wells with PBS to remove non-adherent planktonic cells.
- Biofilm Quantification: Fix the adherent biofilm and stain it with a 1% crystal violet solution for 15-20 minutes. Wash off the excess stain, solubilize the bound crystal violet with 30% acetic acid, and measure the absorbance of the solution spectrophotometrically at 595 nm.
- Calculation of Inhibition: The percent inhibition of biofilm formation is calculated using the formula:
[ (Abs_control - Abs_antibody) / (Abs_control - Abs_blank) ] × 100[31].
Opsonophagocytosis Killing Assay
The functional capacity of antibodies to mediate killing of bacteria is assessed via an opsonic killing assay [18]:
- Preparation: Mix fresh human or mouse neutrophils with bacterial cells (e.g., E. coli ATCC 25922) in the presence of anti-PIA antiserum or control serum. Baby rabbit serum is often added as a source of complement.
- Incubation and Plating: Incubate the mixture under rotation for a set time (e.g., 90 minutes) at 37°C.
- Viability Assessment: After incubation, plate serial dilutions of the mixture onto agar plates. After overnight incubation, enumerate the colony-forming units (CFU).
- Data Analysis: The percentage of bacterial killing is calculated by comparing the CFU from wells with test antiserum to those with control serum. A lethality of 40.48% was reported for anti-PIA antibodies against E. coli [18].
The logical flow from antigen preparation to functional validation is outlined in the diagram below.
Visualization of Cross-Reactive Immunity Mechanism
The mechanism by which anti-PIA antibodies exert cross-reactive effects on E. coli involves binding to the shared PNAG epitope and disrupting key biofilm processes, as illustrated below.
The Scientist's Toolkit: Key Research Reagents and Solutions
Successful research in this field relies on specific biological tools and reagents. The table below catalogues essential materials used in the featured experiments.
Table 2: Key Research Reagent Solutions for PIA/PNAG Biofilm Research
| Reagent / Material | Function and Application in Research | Examples / Specifications |
|---|---|---|
| Bacterial Strains | Source of PIA/PNAG antigen and for functional assays. | S. epidermidis 1457 (PIA+), S. epidermidis M10 (icaA- mutant, PIA-), E. coli ATCC 25922 (biofilm-forming) [18]. |
| Chromatography Media | Purification of native PIA polysaccharide from culture extracts. | Sephacryl S-100 or S-300 for size-exclusion chromatography [31]. |
| Analytical Tools | Confirmation of PIA/PNAG structure and purity. | FTIR Spectroscopy, NMR, colorimetric assays (e.g., MBTH for hexosamine) [18] [31]. |
| Adjuvants | To enhance the immune response during antibody generation in animal models. | 1% Alum [31]. |
| Assay Kits & Components | Quantification of antibody titers and biofilm formation. | ELISA Kits (for mouse IgG), Crystal Violet stain for biofilm quantification, Limulus Amebocyte Lysate (LAL) test for endotoxin detection [31]. |
Discussion and Future Perspectives
The demonstration that anti-staphylococcal PIA antibodies can disrupt E. coli biofilms and promote opsonophagocytosis represents a significant advance in the search for broad-spectrum anti-biofilm strategies. This cross-reactive immunity opens avenues for developing therapeutic antibodies or vaccines targeting a conserved virulence factor across multiple pathogens.
Future research should focus on several key areas:
- In Vivo Efficacy: The promising in vitro and opsonic data require validation in sophisticated animal models of E. coli biofilm-associated infections, such as catheter-associated urinary tract infections or gastrointestinal colonization models [18].
- Human Antibody Development: Exploring the feasibility of humanized or fully human monoclonal antibodies against PIA/PNAG could accelerate translational progress [75].
- Combination Therapies: The potential of anti-PIA/PNAG antibodies to act synergistically with conventional antibiotics or other anti-biofilm agents (e.g., dispersin B, DNase) should be thoroughly investigated [2].
- Strain Coverage and Specificity: Further studies are needed to assess the breadth of activity across clinical isolates of E. coli and other PNAG-producing bacteria, and to rule out potential off-target effects.
In conclusion, within the well-established context of staphylococcal PIA research, the cross-reactive immunity it elicits presents a compelling strategy for combating biofilm-related infections caused by diverse microbial pathogens, holding considerable promise for novel therapeutic interventions in the global fight against antimicrobial resistance.
Polysaccharide intercellular adhesin (PIA), also known as poly-N-acetylglucosamine (PNAG), represents a crucial virulence determinant in staphylococcal biomaterial-centered infections. This whitepaper examines PIA's role as a marker for virulent strains of Staphylococcus epidermidis and Staphylococcus aureus, the leading etiologic agents of implant-related infections. We comprehensively review PIA's biochemical structure, genetic regulation, and functional contributions to biofilm formation and immune evasion. Experimental protocols for PIA detection and quantification are detailed alongside therapeutic approaches targeting this exopolysaccharide. The consolidated evidence positions PIA as both a diagnostic marker for virulent staphylococcal strains and a promising target for novel anti-biofilm strategies.
Implant-associated infections represent a devastating complication in modern medicine, characterized by remarkable therapeutic recalcitrance and recurrence rates exceeding 20% despite aggressive intervention [77]. The success of staphylococci as pathogens in these infections is intrinsically linked to their ability to transition from planktonic to sessile existence through biofilm formation on implant surfaces [2]. Biofilm formation is the main pathogenetic mechanism leading to the chronicity and irreducibility of infections, with bacteria in biofilms resisting antibiotics at concentrations up to 1000 times higher than those effective against planktonic cells [2].
The extracellular matrix of staphylococcal biofilms comprises multiple polymeric substances, among which polysaccharide intercellular adhesin (PIA) has emerged as a particularly critical component and virulence marker [3] [1]. First described in S. epidermidis, PIA is a cationic, partially deacetylated homopolymer of β-1-6-linked N-acetylglucosamine that mediates bacterial accumulation and biofilm formation on biomaterials [3] [2]. Epidemiological studies consistently demonstrate that the presence of PIA biosynthesis genes (ica locus) strongly correlates with infectivity, particularly in device-associated infections caused by S. epidermidis [3].
Structural and Biochemical Characteristics of PIA
Molecular Architecture
PIA is chemically defined as a linear polymer of β-1-6-linked N-acetylglucosamine (GlcNAc) residues with varying degrees of modification [3] [1]. The polymer typically contains approximately 130 residues of N-acetylglucosamine, corresponding to a molecular weight of ~30 kDa, though reported sizes vary from 20 kDa to >460 kDa due to differences in polymerization extent and analytical methodologies [3].
Table 1: Structural Characteristics of Polysaccharide Intercellular Adhesin
| Characteristic | Description | Biological Significance |
|---|---|---|
| Chemical Structure | Linear β-1-6-linked poly-N-acetylglucosamine | Provides structural backbone for biofilm matrix |
| Degree of Deacetylation | 15-20% partially deacetylated | Confers positive charge, enabling interactions with negative surfaces |
| Additional Modifications | ~10% O-succinylation (reports variable) | May influence immune recognition and matrix properties |
| Molecular Weight | ~30 kDa (reports range from 20->460 kDa) | Affects physical properties and potential as vaccine target |
| Net Charge | Cationic | Facilitates attachment to anionic surfaces and biomaterials |
Biosynthetic Pathway
PIA biosynthesis is mediated by the ica (intercellular adhesion) locus, which consists of the biosynthetic operon icaADBC and the regulatory gene icaR [3] [1]. The individual enzymatic components function in a coordinated manner as visualized in the biosynthesis pathway:
The biosynthetic process initiates with IcaA, an N-acetylglucosaminyltransferase that utilizes UDP-N-acetylglucosamine as a substrate to synthesize PIA oligomers [1]. IcaD forms a complex with IcaA, enhancing its catalytic efficiency. IcaC is involved in the externalization and polymerization of the nascent polysaccharide chain on the bacterial surface. Finally, IcaB functions as an N-deacetylase, responsible for the partial deacetylation that confers the cationic properties essential for PIA's adhesive function [1] [2].
Genetic Basis and Regulation of PIA Production
TheicaLocus
The icaADBC operon represents the core genetic machinery for PIA biosynthesis in staphylococci, with significant conservation across species [3]. Presence of the ica locus varies among staphylococcal strains, with epidemiological studies revealing a strong association between ica detection and clinical isolates from device-related infections [3]. In S. epidermidis, the ica genes are present in the A/B cluster at approximately 37%, compared to only 4% in cluster B, suggesting genetic clustering of virulent, biofilm-forming strains [3].
Table 2: Genetic Components of PIA Biosynthesis
| Gene | Function | Phenotype of Mutation |
|---|---|---|
| icaA | N-acetylglucosaminyltransferase; synthesizes PIA oligomers from UDP-N-acetylglucosamine | Abolished PIA production |
| icaD | Enhances catalytic efficiency of IcaA; forms complex with IcaA | Greatly reduced PIA production |
| icaC | Involved in externalization and polymerization of polysaccharide chain | Intracellular accumulation of PIA precursors |
| icaB | N-deacetylase; partially deacetylates PIA (15-20%) | Production of fully acetylated, non-functional PIA |
| icaR | Regulatory gene; represses icaADBC transcription | Constitutive PIA production |
Regulatory Networks
PIA production is strongly influenced by environmental conditions and integrated into global regulatory networks. Two key regulatory systems, agr (accessory gene regulator) and sarA (staphylococcal accessory regulator), play particularly important roles in modulating ica expression and PIA production [77].
The agr quorum sensing system typically functions as a negative regulator of biofilm formation, promoting the dispersal phase that disseminates bacteria to new infection sites [1] [77]. In contrast, sarA enhances biofilm formation through both agr-dependent and agr-independent pathways, with mutation of sarA significantly limiting biofilm formation across diverse clinical isolates [77]. Importantly, spontaneous agr mutants frequently arise in vivo and may promote the transition from acute to chronic infection, whereas spontaneous sarA mutants are rarely isolated from patients, suggesting sarA represents a more stable therapeutic target [77].
Environmental conditions profoundly influence PIA production, with factors such as anaerobiosis, high glucose, increased osmolarity (4% NaCl), and iron availability stimulating ica expression and PIA production [3] [17]. Additionally, staphylococci can undergo phase variation in biofilm production mediated by transposition of insertion sequences in the icaC gene or expansion/contraction of tandem repeats within icaC [1].
PIA as a Virulence Marker in Staphylococcal Infections
Correlation with Clinical Infections
Substantial clinical evidence supports the association between PIA production and staphylococcal virulence in biomaterial-centered infections. Molecular epidemiological studies demonstrate that the presence of ica genes is significantly higher in S. epidermidis isolates from device infection or device-associated blood infection compared to colonization isolates [3]. This correlation establishes PIA as a marker for virulent strains particularly adapted to cause device-related infections.
In S. aureus, while most strains possess ica genes, the reliance on PIA for biofilm formation varies among strains [3] [17]. Some clinical isolates utilize PIA-independent mechanisms for biofilm formation, employing proteinaceous factors instead [1] [17]. This strain-dependent variability underscores the importance of contextualizing PIA as one of several virulence mechanisms in staphylococcal pathogenesis.
Functional Roles in Pathogenesis
PIA contributes to staphylococcal virulence through multiple mechanisms:
- Intercellular Adhesion: PIA mediates bacterial accumulation through direct interactions between the positively charged polysaccharide and negatively charged bacterial surfaces, facilitating multilayer biofilm architecture [3] [1].
- Immune Evasion: The cationic nature of partially deacetylated PIA confers resistance to antimicrobial peptides and protects against phagocytosis by polymorphonuclear neutrophils [3] [71].
- Biomaterial Colonization: PIA enables firm attachment to implanted medical devices, initiating the biofilm life cycle on abiotic surfaces [3] [2].
- Antibiotic Tolerance: Biofilms stabilized by PIA exhibit dramatically enhanced tolerance to conventional antibiotics, contributing to the recalcitrance of device-associated infections [77].
Experimental Methods for PIA Detection and Analysis
PIA Extraction and Purification Protocol
The following standardized method allows for reproducible extraction and purification of PIA from staphylococcal cultures [18]:
Culture Conditions: Inoculate strains in 2 liters of Tryptic Soy Broth (TSB) and incubate at 37°C with gentle agitation (40-50 rpm) for 24 hours to promote PIA production.
Harvesting: Centrifuge cultures at 1000 × g for 20 minutes at 4°C to pellet bacterial cells.
Cell Disruption: Resuspend cell pellets in 20 mL of PBS (pH 7.5) and subject to sonication on ice (four cycles of 30 seconds each).
Clarification: Centrifuge the lysate at 12,000 rpm for 15 minutes to remove cellular debris.
Precipitation: Add trichloroacetic acid (TCA) to the supernatant to a final concentration of 10% (w/v) to precipitate PIA. Incubate overnight at 4°C.
Recovery: Collect precipitate by centrifugation at 10,000 × g for 30 minutes at 4°C.
Dialysis: Resuspend the precipitate in distilled water and dialyze extensively against distilled water to remove residual TCA.
Lyophilization: Lyophilize the purified PIA for long-term storage at -20°C.
Biofilm Formation Assay
The microtiter plate assay represents the standard method for quantifying biofilm formation capacity [18] [17]:
Preparation: Adjust the optical density (OD₆₀₀) of overnight bacterial cultures to 0.7 in BHI broth.
Dilution: Dilute bacterial suspensions 1:200 in BHI broth supplemented with 1% (w/w) D-glucose (BHIGlc) and 4% (w/w) NaCl (BHINaCl) to induce PIA production.
Incubation: Transfer 200 μL of diluted suspensions to polystyrene microtiter plates and incubate for 24 hours at 37°C without agitation.
Washing: Remove planktonic cells by gently washing each well three times with phosphate-buffered saline (PBS).
Staining: Add 150 μL of 1% crystal violet solution to each well and incubate for 15 minutes at room temperature.
Destaining: Remove excess stain and destain with 160 μL of 30% acetic acid solution.
Quantification: Measure absorbance spectrophotometrically at 595 nm. Strains with OD values >0.120 are typically considered biofilm-positive.
Molecular Detection of ica Genes
PCR amplification of ica locus genes provides a reliable method for determining the genetic potential for PIA production. Primers targeting icaA and icaD genes are most commonly used, with amplification conditions optimized for specific staphylococcal species [3] [1]. The simultaneous presence of both genes strongly correlates with functional PIA production and biofilm-forming capacity.
Research Reagent Solutions
Table 3: Essential Research Reagents for PIA Studies
| Reagent/Condition | Function/Application | Experimental Notes |
|---|---|---|
| TSB with 1% Glucose + 4% NaCl | Induction of PIA production under in vitro conditions | Maximizes PIA expression; essential for consistent results |
| Polystyrene Microtiter Plates | Substrate for biofilm formation assays | Surface properties significantly influence attachment |
| Crystal Violet Stain | Biofilm biomass quantification | Standardized washing critical for reproducibility |
| Anti-PIA/PNAG Antibodies | Immunodetection and functional studies | Show cross-reactivity across species (E. coli PNAG) [18] |
| Dispersin B | PIA-degrading enzyme; anti-biofilm studies | Specific glycoside hydrolase targeting β-1,6-GlcNAc linkages |
| S. epidermidis 1457 | PIA-positive reference strain | Wild-type biofilm-forming strain |
| S. epidermidis 1457 M-10/Δica | Isogenic PIA-negative mutant | Essential control for PIA-specific phenotypes |
Therapeutic Implications and Future Directions
PIA as a Therapeutic Target
The critical role of PIA in staphylococcal virulence has stimulated interest in developing therapeutic strategies targeting this exopolysaccharide. Several approaches show promise:
Immunotherapeutic Interventions: PIA has raised considerable interest as a potential vaccine component, with deacetylated PNAG retained on the surface of S. aureus favoring opsonophagocytosis and representing a candidate for immune protection [3] [1]. Antibodies generated against PIA demonstrate significant biofilm inhibition and opsonic activity, with studies showing approximately 40% opsonization-mediated killing of bacterial cells [18].
Enzymatic Disruption: Dispersin B, a glycoside hydrolase that cleaves β-1,6-N-acetylglucosamine linkages, effectively degrades PIA and sensitizes biofilms to antibiotic treatment [2].
Regulatory Interference: Targeting the sarA regulatory pathway presents a promising approach to limiting PIA production without selecting for spontaneous mutants that commonly arise with agr dysfunction [77].
Diagnostic Applications
Detection of PIA or the ica genes provides a valuable diagnostic marker for identifying virulent staphylococcal strains in clinical settings. The strong association between ica presence and device-related infections suggests potential utility in stratifying infection risk and guiding therapeutic decisions [3]. Furthermore, monitoring PIA production could serve as a biomarker for treatment efficacy in biofilm-targeting therapeutic approaches.
Polysaccharide intercellular adhesin represents a defining virulence factor for staphylococci in biomaterial-centered infections. Its well-characterized biosynthesis, regulation, and functional contributions to biofilm formation and immune evasion establish PIA as both a reliable marker for virulent strains and a promising target for novel anti-biofilm strategies. Future research directions should focus on translating this mechanistic understanding into targeted therapeutic and diagnostic applications that address the significant clinical challenge posed by biofilm-associated infections on medical devices.
Conclusion
Polysaccharide intercellular adhesin (PIA) is unequivocally established as a central virulence determinant in staphylococcal biofilm-associated infections, functioning through both mechanical biofilm consolidation and profound immune evasion. The synthesis of knowledge across foundational, methodological, troubleshooting, and validation intents confirms PIA's biosynthetic ica locus as a critical, though not exclusive, mediator of chronic biomaterial infections. Future directions must prioritize the translation of this molecular understanding into clinical applications, including the refinement of PIA-targeted vaccines, the development of biomaterials resistant to PIA-mediated adhesion, and the optimization of quorum-quenching agents for therapeutic biofilm dispersal. The demonstrated cross-reactivity of anti-PIA antibodies with phylogenetically diverse pathogens further underscores the potential for a broad-spectrum therapeutic strategy, positioning PIA research as a pivotal frontier in the ongoing battle against antimicrobial-resistant biofilm infections.
References
- 1. Polysaccharide intercellular adhesin in biofilm [https://pubmed.ncbi.nlm.nih.gov/25713785/]
- 2. Polysaccharide intercellular adhesin in biofilm: structural ... [https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2015.00007/full]
- 3. The staphylococcal exopolysaccharide PIA – Biosynthesis ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7674160/]
- 4. N-Acetylglucosamine [https://en.wikipedia.org/wiki/N-Acetylglucosamine]
- 5. Structural Basis for the De-N-acetylation of Poly-β-1,6-N-acetyl ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4276859/]
- 6. Insights into biofilm architecture and maturation enable ... [https://www.sciencedirect.com/science/article/abs/pii/S2451945624004835]
- 7. Characterization of theN-Acetylglucosaminyltransferase ... [https://www.sciencedirect.com/science/article/pii/S0021925818804653]
- 8. Characterization of the Poly-β-1,6-N-Acetylglucosamine ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3233048/]
- 9. Poly-β-D-(1,6)-N-acetyl-glucosamine (PNAG) glycan ... [https://www.nature.com/articles/s41467-025-61559-7]
- 10. Biofilm Formation, icaADBC Transcription, and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1064907/]
- 11. A Review of Biofilm Formation of Staphylococcus aureus ... [https://www.mdpi.com/2079-6382/12/1/12]
- 12. Correlation between biofilm formation and production of ... [https://www.sciencedirect.com/science/article/abs/pii/S1438422106001196]
- 13. Production of icaADBC-encoded polysaccharide intercellular ... [https://bmcinfectdis.biomedcentral.com/articles/10.1186/1471-2334-10-68]
- 14. Interplay of icaADBC Operon and MSCRAMMs in Biofilm ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9696285/]
- 15. Contribution of icaADBC genes in biofilm production ability of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12084956/]
- 16. Various mutations in icaR, the repressor of the icaADBC ... [https://www.sciencedirect.com/science/article/pii/S1286457924000261]
- 17. Assessment of the biofilm formation capacities ... [https://www.nature.com/articles/s41598-025-00521-5]
- 18. Investigation of the effect of anti-PIA/PNAG antibodies on ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2025.1552670/full]
- 19. Polysaccharide intercellular adhesin or protein factors in ... [https://www.sciencedirect.com/science/article/abs/pii/S014296120601012X]
- 20. The Intercellular Adhesion (ica) Locus Is Present in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC96900/]
- 21. A different path: Revealing the function of staphylococcal ... [https://www.sciencedirect.com/science/article/pii/S0014579314002786]
- 22. Research on the Biofilm Formation of Staphylococcus ... [https://www.mdpi.com/2076-2607/9/7/1534]
- 23. Biology and Regulation of Staphylococcal Biofilm [https://www.mdpi.com/1422-0067/24/6/5218]
- 24. A novel mechanism of phase variation of virulence in ... [https://pubmed.ncbi.nlm.nih.gov/10231490/]
- 25. Phase Variation of Poly-N-Acetylglucosamine Expression in ... [https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1004292]
- 26. Regulation of σB-Dependent Biofilm Formation in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10609180/]
- 27. Purification and Evaluation of Polysaccharide Intercellular ... [https://pubmed.ncbi.nlm.nih.gov/27460584/]
- 28. Polysaccharide intercellular adhesin in biofilm - PubMed Central [https://pmc.ncbi.nlm.nih.gov/articles/PMC4322838/]
- 29. Structures of Streptococcus pneumoniae PiaA and Its Complex with Ferrichrome Reveal Insights into the Substrate Binding and Release of High Affinity Iron Transporters - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3741162/]
- 30. Investigation of the effect of anti-PIA/PNAG antibodies on ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11922938/]
- 31. Evaluation of polysaccharide intercellular adhesion (PIA) and ... [https://bmcresnotes.biomedcentral.com/articles/10.1186/s13104-019-4736-8]
- 32. Application of Spectroscopic Methods for Structural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2885081/]
- 33. Contemporary strategies and approaches for ... [https://www.sciencedirect.com/science/article/pii/S2095177923002794]
- 34. Full Length Article FTIR, 1 H-/ 13 C-NMR spectral ... [https://www.sciencedirect.com/science/article/pii/S2667022424002974]
- 35. A Review of Biofilm Formation of Staphylococcus aureus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9854888/]
- 36. Evaluation of methods to detect in vitro biofilm formation by ... [https://bmcresnotes.biomedcentral.com/articles/10.1186/s13104-018-3820-9]
- 37. Structure, function and contribution of polysaccharide ... [https://pubmed.ncbi.nlm.nih.gov/19913940/]
- 38. Synthesis of conjugated PIA-rSesC and immunological ... [https://pubmed.ncbi.nlm.nih.gov/30990402/]
- 39. Guidelines for the use of flow cytometry and cell sorting in ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9165548/]
- 40. Strategy to combat biofilms: a focus on biofilm dispersal ... [https://www.nature.com/articles/s41522-023-00427-y]
- 41. Enzymatic dispersion of biofilms: An emerging biocatalytic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9478923/]
- 42. The dynamic transcriptome during maturation of biofilms ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2022.882346/full]
- 43. Inhibition of Staphylococcus aureus Biofilm Formation and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9241682/]
- 44. Sticking together: independent evolution of biofilm formation in ... [https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-024-10719-y]
- 45. Protein-based biofilm matrices in Staphylococci - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4261907/]
- 46. Role of Surface Protein SasG in Biofilm Formation by ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2953683/]
- 47. The Intercellular Adhesin Locus Ica Is Present in ... - PubMed [https://pubmed.ncbi.nlm.nih.gov/11388609/]
- 48. The intercellular adhesin locus ica is present in clinical ... [https://link.springer.com/article/10.1007/s430-001-8018-5]
- 49. Presence of the ica operon in clinical isolates ... [https://www.sciencedirect.com/science/article/pii/S1198743X14638507]
- 50. Genetic regulation of the intercellular adhesion locus in ... [https://www.frontiersin.org/journals/cellular-and-infection-microbiology/articles/10.3389/fcimb.2012.00038/full]
- 51. The dual-species biofilm formed by Staphylococcus aureus ... [https://www.nature.com/articles/s41538-025-00581-x]
- 52. Staphylococcus epidermidis Polysaccharide Intercellular ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1082835/]
- 53. Polysaccharide intercellular adhesin (PIA) protects ... [https://pubmed.ncbi.nlm.nih.gov/14764110/]
- 54. Quorum sensing in Staphylococcus infections - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC281656/]
- 55. Quorum sensing in Staphylococcus infections [https://www.jci.org/articles/view/20442]
- 56. Combatting biofilm-mediated infections in clinical settings ... [https://www.sciencedirect.com/science/article/pii/S246823302400015X]
- 57. Quorum-sensing control of biofilm factors in ... [https://pubmed.ncbi.nlm.nih.gov/12934187/]
- 58. Elucidating the Crucial Role of Poly N-Acetylglucosamine from ... [https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0124216]
- 59. Biofilm Formation, c-di-GMP Production, and Antimicrobial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12469266/]
- 60. The Role of Staphylococcal Biofilm on the Surface ... [https://www.mdpi.com/2076-2607/10/10/1909]
- 61. Staphylococcus aureus Biofilm-Associated Infections: Have ... [https://www.mdpi.com/2076-2607/13/4/852]
- 62. Structure, function and contribution of polysaccharide ... [https://www.sciencedirect.com/science/article/abs/pii/S0171933509003197]
- 63. Comparative Transcriptomic Analysis of Staphylococcus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8351120/]
- 64. Comparative transcriptomic analysis of Staphylococcus ... [https://www.sciencedirect.com/science/article/pii/S1438422124000249]
- 65. In vivo gene expression in a Staphylococcus aureus prosthetic ... [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/s12866-016-0695-6]
- 66. Toward an Improved In Vitro Model of Prosthetic Joint ... [https://www.sciencedirect.com/science/article/pii/S2590207525000735]
- 67. In-vitro Investigation of Antibiotics Efficacy Against Uropathogenic... [https://pubmed.ncbi.nlm.nih.gov/32848429/]
- 68. antibiotic tolerance and persistence in bacterial biofilms [https://pmc.ncbi.nlm.nih.gov/articles/PMC6629468/]
- 69. Biofilms: structure, resistance mechanism, emerging ... [https://pubs.rsc.org/en/content/articlehtml/2025/pm/d5pm00094g]
- 70. Immune dysfunction during S. aureus biofilm-associated ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12297603/]
- 71. Extracellular DNA and polysaccharide intercellular adhesin ... [https://www.sciencedirect.com/science/article/pii/S0944501325001326]
- 72. Bacterial Evasion of Host Antimicrobial Peptide Defenses - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC4804471/]
- 73. Effects of polysaccharide intercellular adhesin (PIA) in an ... [https://www.sciencedirect.com/science/article/abs/pii/S1438422115300047]
- 74. An extracellular Staphylococcus epidermidis polysaccharide ... [https://bmcmicrobiol.biomedcentral.com/articles/10.1186/1471-2180-12-76]
- 75. Fighting Staphylococcus aureus biofilms with monoclonal ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6420399/]
- 76. Phenotypic PIA-Dependent Biofilm Production by Clinical ... [https://www.mdpi.com/2076-2615/11/11/3047]
- 77. Staphylococcus aureus Biofilm-Associated Infections [https://pmc.ncbi.nlm.nih.gov/articles/PMC12029350/]